Soós istván – Szabad jános
Kromoszómáink stabilitása
Bármelyikünk élete egy női- és egy hímivarsejt egyesülésével kezdődött. Az első sejt, a zigóta, egy anyai, és egy apai eredetű kromoszómagarnitúrát tartalmaz, 23–23 kromoszómával. Minden kromoszómában egyetlen DNS kettős spirál rejlik, amelyeket nagyon sok, és sokféle fehérjemolekula bugyolál be. A DNS-molekulák részei azok a gének (kb. harmincezer anyai, és ugyanannyi apai eredetű), amelyek meghatározzák öröklődő tulajdonságainkat. Az első sejtből úgy lesz kettő, hogy a DNS-molekulák megkettőződnek, majd miközben a képződő leánykromatidák pakolódnak és tömörödnek, kialakulnak a jól ismert ún. metafázisú kromoszómák, két-két leánykromatidával (1. ábra). A mitózis folyamán a leánykromatidák egyike az egyik, a másik a másik leánysejtbe kerül (2A. ábra). Bár a folyamat egyszerűnek tűnhet, a „mutatványt” nem könnyű úgy megvalósítani, hogy a képződő két leánysejt mindegyikének olyan legyen a kromoszómakészlete, mint az anyasejté volt.

1. ábra. Egy ún. metafázisú
kromoszóma sematikus ábrázolása. A leánykromatidák a centromerjeiknél összetapadnak.
A centromereken épülnek ki a kinetochorok, hozzájuk mikrotubulusok kapcsolódnak,
kinetochoronként 20–40. Forrás: https://www.utexas.edu/courses/
utgeneticstamu/23_38.jpg

2. ábra. A mitózis sematikus ábrázolása. A) Rendes körülmények között egy mitózis eredményeként két, az anyasejttel azonos genetikai állományú leánysejt képződik. Az egymást követő mitózisok eredményeként egyre több ép sejt alkotja a szervkezdeményt. B) Mitotikus nondiszjunkció nyomán olyan sejt képződik, amelyben valamely kromoszómából megszokott kettő helyett három van, valamint egy olyan is, amelyben csupán egy. C) A mitózis során olykor-olykor elvész a kromoszómák némelyike. Bár a szokatlan számú kromoszómát hordozó sejtek többnyire életképtelenek, olykor életben maradnak, osztódnak, és egyre többen lehetnek. A szokatlan számú kromoszómát hordozó sejtek gyakorta daganatok kiindulási forrásai
Sejtjeink megsínylik, ha akár csak egyetlen kromoszómát is „nyernek”, és azt is, ha veszítenek. Vajon milyen mechanizmusok biztosítják a kromoszómák stabilitását az egymást követő sejtosztódások folyamán? Mi történik, ha sérül a „biztosító” mechanizmusok valamelyike, és olyan leánysejtek képződnek, amelyekbe a „rendesnél” több, vagy kevesebb kromoszóma kerül? Mik a meghibásodás következményei? Miként romolhatnak el a „biztosító berendezések”? Mit tehetünk, hogy megelőzzük a bajt? Áttekintésünk a kromoszómastabilitás témakörével ismerteti meg az érdeklődőket.
Testünket mintegy 1014 sejt alkotja. Mindegyik abból az elsőből, a zigótából származik – elmondhatatlanul sok sejtosztódás, mitózis eredményeként –, amely egy petesejt, valamint egy spermium egyesülése nyomán képződött. Tudta ön, hogy a mitózis fontosságát szépen mutatja az a tény is, hogy minden percben milliónyi sejtünk osztódik? A mitózis lényegében olyan folyamat, amely során az anyasejt 46 kromoszómájának két-két összetapadt leánykromatidáját kell előbb elválasztani, majd igazságosan szétosztani a képződő leánysejtek között (2A. ábra). Sejtjeink akkor épek, és láthatják el normálisan funkcióikat, ha bennük – mint annak idején a zigótában – egy anyai, valamint egy apai génkészlet funkcionál. Azok a sejtjeink többnyire elpusztulnak, amelyekben a megszokottnál akár csak eggyel több, vagy eggyel kevesebb kromoszóma van. Mégis, a kromoszómát „nyerő”, vagy a kromoszómát vesztő sejtek olykor életben maradnak. Különösen akkor, ha a „nyert”, vagy az elvesztett kromoszóma rövid, és viszonylag kevés gént tartalmaz. Elfogadott álláspont, hogy sem a kromoszómanyerésnek, sem a -vesztésnek sincs jó vége. Sejtjeink a kromoszóma „nyerését” jobban elviselik, mint a vesztést, és azt is, hogy minél rövidebb a „nyert”, vagy a vesztett kromoszóma, annál nagyobb az esély arra, hogy a sejt életben marad.
A magorsó elkészültét
ellenőrző mechanizmus
Mitózisra készülődve a centromereken
kinetochorok épülnek ki. Egy-egy kinetochorhoz 20–40 ún. kinetochor mikrotubulus
kapcsolódik (1. ábra). Ezek a centroszómából erednek, és alapvető
szerepük van abban, hogy miután a leánykromatidák szétválnak, a centroszóma
felé szállítódnak. Honnan tudják a kinetochorok, hogy melyik irányból érkező
mikrotubulussal kapcsolódjanak? És a mikrotubulusok, hogy melyik kinetochorhoz
tartoznak? Miként jön létre az az állapot, amelyben az egyik kinetochor
minden mikrotubulusa a magorsó egyik pólusába, a másiké pedig a másikba
fut? Ez az állapot a 3. ábrán röviden összegzett események során
alakul ki, és azt eredményezi, hogy kromoszómáink a sejt egyenlítői síkjában
sorakoznak fel (4. ábra).

3. ábra. A kinetochorok
és a kinetochormikrotubulusok
közötti kapcsolat időben
történő fontosabb mozzanatai vázlatosan. Az, hogy a kromoszómák a sejt
egyenlítői síkjában sorakoznak fel azoknak az
erőknek köszönhető, amelyekkel
a mikrotubulusok húzzák a kinetochorokat. Forrás: Tanaka and Desai, Curr
Opin Cell Biol 20, 53, 2008.

4. ábra. A mitózis folyamán
az ún. metafázisú kromoszómák, miközben
mikrotubulusokkal kapcsolódnak,
a sejt egyenlítői síkjában sorakoznak fel. Forrás: https://wikispaces.psu.edu/display/230/Mitosis,
+Cell+Division+and+the+Cell+Cycle
Nos, sejtjeinkben funkcionál egy, a magorsó elkészültét ellenőrző mechanizmus (angolul: spindle assembly checkpoint), amely addig nem engedi tovább a mitózis eseményeit, ameddig a sejt nem készült fel a folytatásra. Vagyis: (i) minden kinetochorhoz kapcsolódnak mikrotubulusok, (ii) valamely kinetochor összes mikrotubulusa ugyanahhoz a centroszómához fut, és (iii) a leánykromatidák kinetochorjainak egyike az egyik, másika a másik centroszómához kapcsolódik. E feltételek teljesülése alapvetően fontos ahhoz, hogy a leánysejtek kromoszómakészlete az anyasejtével azonos legyen (5. ábra).

5. ábra. A leánykromatidák
szétválásának mechanizmusa. A metafázist megelőzően a kinetochorokhoz olyan
fehérjeféleségek kapcsolódnak, mint a Mad2 és BubR1. Ezek a fehérjék minden
olyan kinetochorban aktívak, amelyek nem, vagy csak tökéletlenül kapcsolódnak
mikrotubulusokkal. Az aktív Mad2- és BubR1-fehérjék gátolják az APC/CCdc20
(angolul: anaphase-promoting complex/cyclosome) komplex aktiválódását.
A Mad2- és BubR1-fehérjék addig aktívak, és a mitózis eseményei nem léphetnek
tovább, amíg nem áll mindegyik kromoszóma mindkét kinetochorja készen,
kapcsolódik egyikük minden mikrotubulusával az egyik, másikuk a másik centroszómához
(4. ábra). Amennyiben a magorsó kialakulását ellenőrző mechanizmus mindent
rendben talál, megszűnik a Mad2/BubR1-gátlás, aktiválódik az APC/CCdc20
komplex, és lebontja a szekurin, valamint a Ciklin B-molekulákat. A szekurin
a szeparáz gátlója. Miután a szekurin lebomlik, aktiválódnak a szeparáz
enzim molekulák, elbontják a kohezin komplexeket. A kohezin komplexek tartották
együtt a leánykromatidákat. A kohezin komplexek lebomlása után szabaddá
válnak a leánykromatidák, és mint önálló kromoszómák kezdhetnek szállítódni
a magorsó pólusai felé. Forrás: https://molbiol4masters.masters.grkraj.org/html/
Cell_Cycle_And_Its_Regulation.htm
Mitotikus katasztrófa
Bár a magorsó elkészültét
ellenőrző mechanizmus meglehetősen megbízhatóan látja el feladatát, olykor-olykor
hibázik. Megtörténik, hogy az egyik leánysejtbe a megszokottnál eggyel
több, a másikba eggyel kevesebb kromoszóma kerül (2B. ábra). Ezekben
az ún. aneuploid sejtekben egy következő „biztosító” rendszer, az ún. mitotikus
katasztrófa aktiválódik azzal a feladattal, hogy az aneuploid sejt a lehető
leghamarabb elpusztuljon. Eleddig pontosan nem ismert jel(ek) alapján a
sejt mintegy „észleli”, hogy baj van, és a legegyszerűbb megoldást választva
igyekszik elpusztítani magát: a programozott sejthalállal (apoptózis),
vagy nekrózissal. A két folyamat részleteire itt nem térünk ki. A mitotikus
katasztrófa egyik ismert megoldása az, hogy a szokatlan számú kromoszómát
(vagy kijavíthatatlanul károsodott DNS-t) tartalmazó sejtmagból olyan molekulák
szabadulnak ki, amelyek inaktiválják a centroszómákat, miáltal azok nem
szervezhetnek több sejtosztódást. Vagyis szervezetünk – a mitotikus katasztrófa
révén – képes „kigyomlálni” azokat a sejteket, amelyek létünket veszélyeztethetik.
Az elhalt sejteket pedig az épek pótolhatják. A „kromoszómastabilitás”
(amit gyakran genomstabilitásnak is neveznek) lényegében azt jelenti, hogy
a sejtek az egymást követő mitózisok folyamában képesek megőrizni a rájuk
jellemző ép kromoszómakészletet. A kromoszómastabilitás megőrzésének az
is része, hogy a szervezetek a szokatlan számú kromoszómát hordozó sejtjeiktől
megszabadulnak.
Túlélő aneuploid sejtek
Bár mind a magorsó elkészültét
ellenőrző mechanizmus, mind a mitotikus katasztrófa meglehetősen pontosan
teljesíti feladatát, meg-megesik, hogy az aneuploid sejtek némelyike életben
marad. Ezek a sejtek osztódhatnak, szokatlan tulajdonságukat átörökíthetik
utódsejtjeikre. Lényegében genetikai mozaikok képződnek (2B. és 2C. ábra),
olyan élőlények, amelyek testét eltérő genetikai tartalmú sejtek alkotják.
Nyilvánvaló, hogy minél több mitózis játszódik le az aneuploid sejt képződését
követően, annál nagyobb lesz a mozaik folt.

6 ábra. Életvonalak egy
Downmozaik
ember két tenyerén. Amíg
a bal tenyéren az életvonalak lefutása
normális, a jobb kézen
– a Downszindrómásokéra
jellemzően – csak egyetlen
életvonal képződött.
Forrás: https://www.pyroenergen.com/
articles07/images/simian-line.jpg
Az említett mozaikosságra két példát említünk. (1) Az ún. Down-mozaikos embertársaink testét kétféle sejt alkotja. Sejtjeik zömében két, kisebbik hányadában három 21. kromoszóma van (6. ábra). Ez az állapot kedvez az Alzheimer-kór, valamiknt az akut leukémia kialakulásának. (2) Azok a sejtek, amelyekben a megszokott kettő helyet csupán egy van a 22. kromoszómából, gyakorta meningeomák kiindulási helyei (7. ábra).

7. ábra. A csak egyetlen
22. kromoszómát
hordozó sejtek meningeoma
daganat
forrásai lehetnek. Forrás:
https://
patikapedia.hu/meningeoma
Ma már sok olyan daganatféleséget ismerünk, amelyek sejtjeiben sem a magorsó elkészültét ellenőrző, sem a mitotikus katasztrófa mechanizmusa nem működik. Érthető, hogy az ilyen sejtek utódsejtjeiben a legkülönfélébb kromoszóma-garnitúrájúak fordulnak elő. A kapcsolat az aneuploidia és rákos állapot között annyira megalapozott, hogy az aneuploid sejtek létét a rákos állapot jelének fogadják el.
Miként okoz bajt az extra
kromoszóma, vagy a kromoszóma hiánya?
A Down-szindrómás embertársaink
sejtjeiben, mint az imént említettük, három 21. kromoszóma van. DNS-ük
vizsgálata alapján arra derült fény, hogy mindegyiknek minden (kromoszómánként
kb. 300) génje tökéletesen ép. Akkor vajon miért alakulnak ki a Down-szindróma
jellegzetes tünetei? A magyarázat minden bizonnyal az, hogy sejtjeinkben
nem a DNS, hanem az általa kódolt fehérjemolekulák látják el a rájuk jellemző
feladatokat. Akkor, ha koncentrációjuk megfelelő a sejtekben. Nos, génjeink
zömének kifejeződése szabályozott: az általuk kódolt fehérjék koncentrációja
a sejtben független a kódoló gének számától. Vannak azonban olyan génjeink
is, amelyek kifejeződése nem szabályozott: a fehérje koncentrációja függ
a kódoló gének számától. Ha például a sejtben valamely ilyen génből nem
kettő, hanem három van, a kódolt fehérje koncentrációja 50%-al meghaladja
a normális szintet. Ha pedig csak egy, úgy a normális felére csökken. A
meghibásodott sejtfunkció oka tehát az, hogy az aneuploid sejtekben némely
fehérjeféleségek koncentrációja felborult: megnövekedett, vagy csökkent.
Például a DYRK1A gén, dózisának 50%-os növekedése felelős a Down-szindrómás
embertársaink csökkent tanulóképességéért, gyenge emlékezőtehetségéért,
szellemi fogyatékosságáért.
Hibák a mitózis során
A mitózis során bekövetkező
hibák vagy genetikai, vagy környezeti eredetűek. Minthogy mind a magorsó
elkészültét ellenőrző, mind a mitotikus katasztrófa szereplői fehérjemolekulák,
őket gének kódolják. Várható tehát hogy, amennyiben a „biztosító” folyamatok
bármelyikében szereplő gének valamelyike sérült, csökken a kromoszómák
stabilitása, és a megszokottnál gyakrabban képződhetnek aneuploid sejtek.
Az aneuploid sejtek aztán sokféle baj forrásai. A genetikai okok tárháza
bőséges. (i) Sem az anyai, sem az apai eredetű gén nem, vagy csak alig
funkcionál. Az ilyen esetekben általában már a gyermek, vagy fiatal felnőttkorban
daganatos betegségek alakulnak ki. (ii) Lehetséges, hogy valaki egyik szülőjétől
a gén ép allélját örökli, a másiktól pedig olyat, amely nem, vagy csak
alig-alig funkcionál. Az ilyen emberek sejtjeiben hatféle (itt fel nem
sorolt) olyan genetikai esemény bármelyike bekövetkezhet, amelyek nyomán
sejtjeinek egyike-másika elveszti az ép gént, és kész a baj. (iii) Előfordul,
hogy testünk valamely sejtjében – mutációk nyomán – az anyai, és az apai
eredetű ép gén is elveszti funkcióját. (Nem egyszerre, hanem csak hosszú
idő elteltével.) (iv) Az is meg-megtörténik, hogy a képződő mutáció ún.
domináns negatív természetű: terméke megakadályozza az ép géntermék normális
funkcióját. Az említett négy példa eredménye ugyanaz: csökken kromoszómáink
stabilitása, és elkezdődhetnek a kellemetlen folyamatok. Példaként a BubR1
gént említjük, melynek terméke a kinetochorhoz kapcsolódik, és szereplője
a magorsó elkészültét ellenőrző mechanizmusnak. Frio és munkatársai (New
Engl J Med 30, 363, 2010) közölték, hogy egy 34. éves emberben, akinek
sejtjeiben sem az anyai, sem pedig az apai eredetű BubR1 gén nem funkcionált,
gyakran képződtek aneuploid sejtek, amelyek egyikéből-másikából végül vastagbél-
és gyomorrák alakult ki.
Léteznek olyan környezeti tényezők is, amelyek úgy eredményezik az aneuploid állapot létrejöttét, hogy akadályozzák az említett két mechanizmus valamelyikének normális funkcióját. Például a vinkrisztin és a vinblasztin (a rózsameténg, Cataranthus roseus) alkaloidái úgy okoznak mitotikus katasztrófát, hogy a tubulinokhoz kötődve tönkreteszik a mikrotubulusokat és a magorsót (3. ábra). Ezért is használják a vinkrisztint és a vinblasztint a féktelenül osztódó daganatos sejtek elpusztítására. Igen ám, csakhogy a kinetochor mikrotubulusokra gyakorolt hatásuk révén zavarják a magorsó elkészültét ellenőrző mechanizmust is az ép sejtekben, ami pedig nem kívánatos „mellékhatás”. Hasznos lenne tehát olyan megoldásokat kidolgozni, amelyekkel meg lehet vizsgálni: miként befolyásolják a különféle kémiai, fizikai és biológia hatások a kromoszómák stabilitását, azaz indukálják kromoszómák nyerését és/vagy vesztését. A feladatot indokolja, hogy bizonyított a kapcsolat a szerves oldószerek használata, a rendellenes lefolyású mitózisok és a rákos daganatok kialakulása között, például a szobafestők, a cipészek, vagy a karosszéria lakatosok életében.
Kromoszómavesztés kimutatása
Intézetünkben olyan genetikai
módszert dolgoztunk ki a kromoszómavesztés kimutatására, amely egyszerű,
érzékeny, kvantitatív, meglehetősen gyors, és olcsó is. Alapja az, hogy
a muslicasejtek életképessége nem csökken, miután elvész belőlük az Y-kromoszóma.
(Az X0-, Y-kromoszóma nélküli muslica hímek tökéletesen életképesek, bár
sterilek. Az Y-kromoszóma tucatnyi génjének zöme a spermiumok mozgásához
szükséges.) A többi kromoszóma bármelyikének vesztése nyomán a sejtek elpusztulnak,
illetve életképességük erősen csökken. Ahhoz, hogy az Y-kromoszóma vesztését
észre tudjuk venni, egy olyan Y-kromoszómát konstruáltunk, amelybe beültettük
az mwh+ gént. Itt mwh+ az mwh (multiple wing hairs) marker-mutáció ép génváltozatát
jelöli. A muslica mwh/mwh szárnysejtjei a szokásos sejtenkénti egyetlen
szőröcske helyett egy egész kis csokorra valót fejlesztenek (8. ábra).
(Az mwh mutáció a muslica harmadik kromoszómájához kapcsoltan öröklődik.)
Amennyiben az mwh/mwh sejtek tartalmazzák az mwh+Y kromoszómát – benne
az ép mwh+ gént –, csak egyetlen szőrszálat növesztenek, ám ha elvesztik,
akár ötöt is. Ha tehát valamilyen hatás nyomán valamely sejtből elvész
az mwh+Y kromoszóma, a képződő X0-sejt és leszármazottai a szokásos sejtenkénti
egyetlen szőrszál helyett többet képeznek, a szárnyon egy mozaik folt alakul
ki. A gondolatmenet megfordítható: az X/mwh+Y; mwh/mwh hímek szárnyain
képződő mwh mozaik foltok jelenléte azt mutatja, hogy a vizsgált genetikai,
vagy környezeti hatás kromoszóma-instabilitást, -vesztést okoz. Ha tehát
valamilyen kezelés nyomán az mwh mozaik foltok gyakorisága meghaladja a
kontroll értékét, a kezelés kromoszóma-instabilitást okoz.

8. ábra. Az mwh+Y-kromoszóma – és vele együtt az ép mwh+ gén – vesztése nyomán képződő leánysejt, majd leszármazottai életben maradnak, és a szokásos sejtenként egyetlen szőr helyett egész kis csokorra valót képeznek. A szaggatott fehér vonal egy mwh mozaikfoltot kerít körül, benne két mwh/mwh sejttel
A szárnykezdemények az embriógenezis folyamán alakulnak ki, egyenként 30–50 sejtből. A sejtek a lárvaállapotban tízóránként osztódnak, miközben számuk 10–11 sejtciklus során ezerszeresére, nagyjából ötvenezerre nő (9. ábra). A kifejlett szárnykezdemény egyetlen sejtréteg, amelynek mintegy harmincezer sejtje alkotja majdan a kifejlett szárnyat és képez sejtenként egyetlen szőrszálacskát. Az mwh marker-mutáció az mwh/mwh sejteket észrevehetővé teszi, még akkor is, ha harmincezer között csak egyetlen egy mwh/mwh.
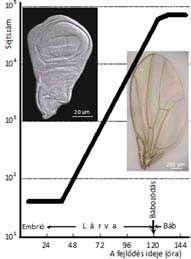
9. ábra. A muslica szárnykezdemény
sejtjei a lárva életszakaszban tíz
óránként osztódnak. Számuk
exponenciálisan gyarapszik a
bábozódásig. A bábállapot
végén a kezdemény sejtjeinek kb. feléből egy
szárny képződik. Az ábra
baloldala egy kifejlett szárnykezdemény, a jobb oldala egy szárny képét
mutatja
A kromoszómavesztések gyakoriságát a következő megfontolások alapján lehet „megmérni”. Amennyiben egy mozaik foltot m sejt alkot, úgy kijelenthetjük, hogy az mwh+Y kromoszóma vesztésének időpontjában a szárnykezdemény C/2m sejtből állt. (Tudvalevő, hogy a szárnykezdemény sejtjei nagyjából azonos mértékben járulnak hozzá a szárnyat alkotó sejtek képzéséhez. Itt C az egy szárnyat alkotó összes sejt száma, 30000. Az mwh mozaik foltot alkotó sejtek számát kettővel szorozni kell, mert az mwh+Y kromoszóma vesztését követően az mwh marker-mutáció csak az egyik utódsejtet jelöli meg; 8. ábra.) Ha N szárnyat vizsgálunk, a kezelésnek kitett sejtek száma N C/2m. Minthogy ugyanannyi sejtből képződik mwh mozaik folt, ahányból elvész az mwh+Y kromoszóma, a kromoszómaveszés gyakoriságát, erősségét az f = n 2m/N C öszszefüggéssel lehet jellemezni. (n az mwh mozaik foltok száma N szárnyon.)
A kísérlet úgy történik, hogy olyan X/mwh+Y; mwh/mwh lárvákat teszünk ki valamely vizsgálandó hatásnak, amelyekben a szárnykezdemények mindegyike néhány ezer sejtből áll. Vagy úgy, hogy a tanulmányozott vegyületet a táptalajba keverjük, amit a lárvák elfogyasztanak, vagy úgy, hogy szerves oldószerek gőzében tartjuk a lárvákat, esetleg úgy, hogy vékony tűvel beléjük injektáljuk. A lárvákból négy-öt nap multán kifejlődő muslicák szárnyait lecsippentjük, és mikroszkópban kb. 400-szoros nagyítással vizsgáljuk. Feljegyezzük az mwh mozaik foltok számát (n), méretét (m), a szárnyak számát (N), hogy aztán meghatározzuk az mwh+Y kromoszóma vesztésének gyakoriságát. Nos, a spontán mwh+Y kromoszómavesztés gyakorisága sejtosztódásonként mindössze 1,7 x 10–4, és hússzorosára nő, ha a táptalaj 1 mg/ml kolhicint tartalmaz.
Minthogy az alapvető sejtfunkciók, a bennük szereplő gének és termékeik az evolúció során erősen megőrződtek, módfelett valószínű, hogy azok a hatások, amelyek instabillá teszik a muslica mwh+Y kromoszómáját, a többire is ugyanilyen hatással vannak. És nemcsak a muslica, hanem a magasabb rendű élőlények – köztük az ember – kromoszómáira is. Lényegében tehát az mwh+Y-kromoszómával, a foltos muslica szárnyak alapján egynémely emberi nyomorúságnak vehetjük elejét.
Az írás az OTKA NI 69180
számú pályázata alapján készült.
| Természet Világa, | 143. évfolyam, 9. szám,
2012. szeptember
https://www.termvil.hu/ https://www.chemonet.hu/TermVil/ |