A mikrobák védelmében
Kulcsszereplők és segítőtársak
Nem kétséges, hogy a XIX–XX. században megváltozott tisztálkodási–tisztasági szokások jelentősen hozzájárultak ahhoz, hogy a Föld fejlettebb részein megszűntek a korábban hatalmas pusztítást okozó járványok. Talán éppen ezért általános az a nézet napjainkban is, hogy a baktériumok és általában a mikrobák veszélyesek, védekezni kell ellenük, irtani kell őket például úgy, hogy antibakteriális szappannal mosakszunk.
A valóságban a mikrobiális és a magasabb rendű élőlények között nemcsak ellentétes érdekű gazda–parazita, hanem legalább olyan sokrétű, azonos érdekű mutualista kapcsolat is van. Írásomban néhány, a bioszféra működésében kulcsszerepet játszó vagy különösen érdekes mikroba–gazda kapcsolatot mutatok be, majd az ember (és az összes emlős) emésztőrendszerében élő mikrobaközösség működéséről és fontosságáról is szólok. Mindezen kérdéseket elsősorban evolúciós ökológiai szempontból vizsgálom: azaz elsősorban arra a kérdésre keresem a választ, hogy milyen mechanizmusok miatt stabilak evolúciósan ezek a kölcsönhatások.
Baktériumok kulcsszerepben
Minden élőlénynek szüksége
van nitrogénre, mivel az amino- és nukleinsavak (a fehérjék, az RNS és
DNS alkotókövei) nitrogént tartalmaznak. A nitrogén óriási mennyiségben
áll rendelkezésre a levegőben, azonban az ott lévő N2-molekulák
nagyon nehezen lépnek kémiai reakcióba. A nitrogén biológiai megkötése
során egy enzim segítségével és energia felhasználásával az N2-molekulából
ammónia (NH3) keletkezik. Ezt a kulcsfontosságú reakciót azonban
csakis egyes baktériumok tudják végrehajtani. Az ammóniát ezután más baktériumok
ammóniummá (NH4+), nitritté (NO2–), majd
nitráttá (NO3–) alakítják, így lesz felvehető a nitrogén
a legtöbb növény számára. Sivár egy hely lenne tehát a Föld a nitrogénkötő
cianobaktériumok, rhizobiumbaktériumok vagy zöld kénbaktériumok nélkül.
Az elpusztult szerves anyag lebontását is elsősorban baktériumok és gombák
végzik. A lebontás során többek között az elhalt élőlényekbe beépült szerves
nitrogént alakítják vissza ammóniummolekulákká. Azaz a mikróbák nemcsak
a nitrogén megkötésében, hanem annak körforgásában is elengedhetetlenül
fontosak (1. ábra).
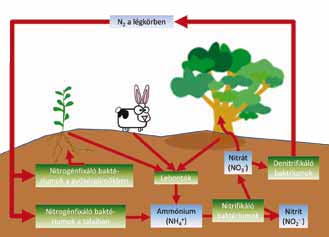
1. ábra. A szárazföldi
biotikus nitrogénciklus vázlatosan. A
ciklus összes lépésében
a baktériumoknak kulcsszerepük van
A nitrogénkötő baktériumok között különösen érdekesek a rhizobiumbaktériumok. Ezek a baktériumok a pillangósvirágú növények speciálisan kialakított gyökérgümőiben élve képesek a nitrogén megkötésére és ammónium előállítására. A növény cserébe szénhidrátokkal, aminosavakkal és fehérjékkel látja el a baktériumot. Amikor a magból egy új palánta kel ki, a gyökere még nem tartalmaz baktériumokkal megrakott gümőket. A gümők kialakulásához először elkezd a gazda a baktériumokkal „beszélgetni”. Kémiai jeleket bocsát ki (flavonoidok formájában). Ezen jelek hatására a közelben lévő baktériumok elindulnak a jel forrása felé, és közben speciális gümőképző molekulákat bocsátanak ki. A gümőképző molekulák hatására a növény hajszálgyökereinek szerkezetében változások jönnek létre, melyek lehetővé teszik, hogy a baktériumok be tudnak jutni ide, s a gümőképződés beindul. A harmónia tökéletesnek tűnik, azonban e növény–baktérium kapcsolat evolúciós stabilitása mégsem érthető meg könnyen.
Együttműködők és csalók
A baktériumok (és általában
az egysejtűek) esetében igen gyakori, hogy a sejten kívüli térbe juttatnak
valamilyen, mindenki számára elérhető tápanyagot vagy enzimet, esetleg
antibakteriális molekulát, ami a környéken lévő minden hasonló egyed számára
elérhető. Ilyenkor azok a változatok, melyek nem vesznek részt a javak
termelésében, a termelőknél több energiát fordíthatnak a szaporodásra vagy
a túlélésre, ezért várhatóan kiszorítják a termelőket a populációból.
A valóságban azonban nem ezt tapasztaljuk. Kérdés, hogy mi korlátozza a
csalók előretörését?
Visszatérve a korábban bemutatott példánkhoz, képzeljük el, hogy a növény hajszálgyökereiben elindult a gümőképződés, és a gümőkben elkezdenek szaporodni a baktériumok. De mi történik akkor, ha olyan baktériumok is bejutnak a gümőbe, melyek nem szolgáltatnak ammóniumot a növénynek, de aktívan kihasználják a gazda nyújtotta előnyöket? Mivel nem költenek az ammónium előállítására, gyorsabban fognak szaporodni, mint a mutualista baktériumok. Korábbi okfejtésünk alapján, ezek a csaló törzsek előbb-utóbb teljesen kiszorítják az együttműködő törzseket, a növény–baktérium mutualizmus megszűnik. Azonban több mint 19 000 pillangós virágú növényfajt és legalább 90–100, ezek gyökérgümőiben élő baktériumfajt ismerünk, ráadásul ez a mutualista kapcsolat körülbelül 60 millió éve stabilan jelen van szinte az egész Földön. Akkor hol a hiba a fenti okoskodásban, azaz milyen evolúciós mechanizmusok tartják fenn ezt a mutualista kapcsolatot?
Érdekes módon nem attól vagyunk zavarban, hogy semmi jó ötletünk sincs, amivel megmagyarázhatnánk a csalók kordában tartását, hanem inkább az a gond, hogy számos, a mutualista kapcsolatot fenntartó hipotézist ismerünk. Az egyik magyarázat szerint a gazda még a baktérium felvétele előtt szelektálni tud a számára „jó” és a „rossz” baktériumok között. Ezzel rokon az az elképzelés, amely szerint a gazdában olyan körülmények alakultak ki az evolúció során, amelyek csak a mutualisták számára kedvezőek. Így „szűri ki” a gazda a számára megfelelő partnereket. Az is elképzelhető, hogy a gazda nem tud a kapcsolat létrejötte előtt szelektálni a baktériumok között, de érzékeli, ha egy gümőben rosszul mennek a dolgok, s arra reagál valahogy. Például az ammónium csökkenése a gümőben az anyagcserét lassítja, így az olyan gümőkben, ahol a csalók túl sokan vannak, a gümő fejlődése lassul vagy le is állhat. Van tehát egy automatikus visszacsatolás, mely bünteti a csalókat. Ehhez közel áll az az elképzelés, amely szerint szelekciós hatások miatt ez a visszacsatolás nem automatikus, hanem éppen azért jött létre, hogy a csalókat büntesse. Nyilván kísérletileg nagyon nehéz az automatikus és a szelekció okozta büntető viselkedést szétválasztani. Jelenleg annyit tudunk, hogy vannak olyan pillangósvirágúak, ahol kísérleti körülmények között a mesterségesen csökkentett ammóniatermelés csökkenő gümőméretet okoz.
A közelmúltban Marco Archettivel közösen kidolgozott elméletünk szerint a csalók általában akkor sem tudják a mutualistákat kiszorítani, ha a növény semmilyen válogatással vagy büntetéssel nem védekezik. Ennek oka az, hogy a mutualisták termelte ammónium mennyisége és a baktériumok által érzékelt haszon egy telítődő görbe szerint alakul (2. ábra). Amennyiben még az is igaz, hogy a termelt javak okozta nyereség a termelő környezetében kicsit erősebben érvényesül, mint tőle távolabb, akkor a termelők és a csalók stabil együttélése az egyik tipikus egyensúlyi helyzet. Mindent összevetve, egyelőre nem tudjuk pontosan, hogy a gazdanövény hogyan akadályozza meg a csaló baktériumok terjedését. Sokféle mechanizmus működhet a probléma megoldására.

2. ábra. A baktériumok
által érzékelt nyereség telítődik az
együttműködő egyedek
gyakoriságának növelésével. Kezdetben
egyre gyorsabb, majd
egy pont után egyre lassabb a növekedés.
A termelők (piros) és
a csalók (fekete) nyereségfüggvénye különböző.
A termelés költséges,
ezért van a piros görbe kezdetben a
fekete alatt, de a termelők
átlagosan kicsit nagyobb nyereséget
érzékelnek, ezért gyorsabban
nő a nyereségük, s persze előbb is
telítődik. Van egy instabil
egyensúlyi pont (fehér kör), mely fölött
a termelők nyeresége
nagyobb, és egy stabil egyensúlyi pont,
mely fölött a csalók
nyeresége a nagyobb. Ha az együttműködők
gyakorisága kisebb, mint
az instabil pontban, akkor a versenyt
a csalók nyerik egyébként
stabilan együtt élnek a csalók és a termelők.
Az ábra tetején a nyilak
a szelekció irányát, a fehér és fekete
körök rendre az instabil
és a stabil állapotokat jelölik
És a gazdanövény miért nem csapja be a baktériumot, kérdezhetnénk? Mert nem tudja! Ugyanis egy csaló gazda, aki nem vagy alig szolgáltat szénhidrátokat és aminosavakat a baktériumoknak, az a saját baktériumait sanyargatja. Így a csaló gazdanövény rosszabbul fejlődik, mint a mutualista növények, tehát kevesebb (és) vagy gyengébb utódai lesznek, így ez a típus előbb vagy utóbb kiszorul a populációból.
Érdekes baktériumok, különleges
állatok
Ebben a fejezetben olyan
baktériumok és állatok közötti kölcsönösen előnyös kölcsönhatásokról szólok,
melyek meglepőek és talán kevéssé ismertek, ráadásul az evolúciós magyarázatuk
is kihívást jelent.
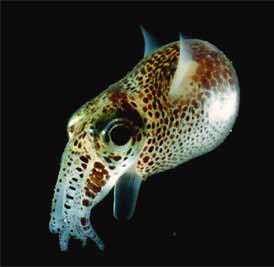
3. ábra. A hawaii kurtafarkú
tintahal éjjeli pompájában.
A kifejlett állat 3–5
cm hosszú. A baktériumok
fényét az állat lencsékkel,
tükrökkel és színszűrővel
állítja be a kívánt irányúra
és színűre (Forrás:
https://www.devbio.biology.gatech.
edu/?page_id=303)
Világítás baktériumok
segítségével
A kurtafarkú tintahalak
éjszakánként halvány fénnyel világítanak, nappal pedig elbújnak a tengerfenéken
(3.
ábra). A legtöbbet a hawaii kurtafarkú tintahal (Euprymnascolopes)
biológiájáról tudunk, megállapításaink ezért ezen a fajon végzett kísérleteken
alapulnak. A tintahal a felszíni vizekben táplálkozik éjjelente, de ilyenkor
egyben lehetséges táplálék is a ragadozó halak számára. Ha a Hold
szépen világít a hawaii égbolton, akkor annak a fénye éppen
elég, hogy a ragadozók könnyen felfedezzék a vízben úszkáló tintahalakat.
Pontosabban könnyen felfedeznék, ha a tintahal által kibocsátott halvány,
lefelé irányuló fény nem akadályozná meg, hogy a tintahalat az árnyékuk
láthatóvá tegye. A jelenség érdekessége, hogy a tintahalak fényét a speciális
világítószervükben élő Vibrio fisheri baktériumok adják. Az
ikrákból kikelő ivadékokban még nem található meg ez a baktérium, hanem
életük e korai fázisában a tengervízből veszik fel őket. De hogyan válogatja
ki a tintahal éppen a V. fisheri-t a vízben lebegő többi száz meg
száz faj egyedei közül? A folyamat – némileg elnagyolva – a következő:
a fényképző szerv külső felületén egy speciális nyák jelenik meg, és csillós
karok nőnek, melyek a vízben lévő baktériumokat a nyákhoz hajtják. A baktériumok
hatására a nyákos felületből kiindulva vékony járatok alakulnak ki, melyeken
keresztül a baktériumok a fényképző szervbe jutnak. Valószínűleg a nyákban
is van előszűrés, azaz csak bizonyos baktériumok számára kedvező ez a környezet.
Ezután a nyák és a járatok megszűnnek, az állat kívülről többé nem vesz
fel baktériumokat. A fényképző szerv azonban nem kellemes hely a legtöbb
baktérium számára, mivel magas koncentrációban tartalmaz hidrogén-peroxidázt
(H2O2). A V. fisheri kivétel ilyen szempontból, mert
ennek a baktériumnak van egy speciális luciferáz (oh, Lucifer, a fényhozó)
nevű enzime, mely a veszélyes oxigénionokat fogyasztja, s közben fény formájában
távozik a felszabaduló energia. A gazdaállat „fertőtlenítőjét” csak a V.
fisheri baktériumok élik túl, s mellékesen világítást szolgáltatnak a gazdának.
A gazda a fertőtlenítőszer mellett tápanyagban gazdag környezetet is biztosít
a baktériumok számára, ahol azok gyorsan szaporodnak. Reggelente ugyanakkor
meg is szabadul a baktériumok 90–95%-tól, hogy estére megint egy optimális
méretű populáció lakja a fényképző szervét.
Akkurátus mezőgazdák és
növényvédő szereik
A mezőgazdaság úgy 20 000
évvel ezelőtt kezdődött az erdei irtásgazdálkodással, s fejlődése tart
egészen napjaink génmódosított állat- és növénytermesztéséig vagy biogazdálkodásáig.
Azonban a mezőgazdaságot a levélvágó hangyák már jóval korábban, kb. 50–60
millió éve elkezdték alkalmazni. A levélvágó hangyák a begyűjtött levéldarabkákat
kolóniájukban gyűjtik. A hangya a nagymennyiségű cellulózt tartalmazó leveleket
nem tudja megemészteni, abban egy, a hangyafajjal együtt élő gombafaj segít
neki. A begyűjtött levelek és a föld alatti nedves klíma kitűnő élőhely
a gomba számára, ahol az gyorsan szaporodik. A hangyák a gombák zsenge
termőtesteit fogyasztják, a kapcsolat mindkét fél számára előnyös.
De mi történik, ha kártevő megtámadja meg a kertészetét? A hangyák
igazi biogazdák: egyes kasztok a telep tisztántartásával, mások a hangya
által felismert kórokozó gombák eltávolításával foglalkoznak. Vannak azonban
mikrogombák és baktériumok is, melyek megfertőzhetik a gombaültetvényt,
ezek ellen baktériumokkal védekezik a hangya. A hangyák kültakaróján
olyan baktériumok élnek, melyek különböző antibiotikumokat termelnek. A
baktériumok megtelepedését speciális mélyedések segítik, melyeket a hangya
kutikulája alatt elhelyezkedő mirigyekből táplál. A hangya a gombaültetvényben
sürgölődve szétkeni az baktériumok által termelt „antibiotikum-koktélt”,
így akadályozva meg a kórokozók terjedését Az új kolónia alapításakor
az alapító királynő a szájüregében lévő speciális zsebbe gyűjt mintát a
korábbi kolónia gombájából, így biztosítja, hogy az új kolónia a kedvező
gombával éljen együtt. Azonban arra nincs egyértelmű bizonyíték, hogy a
korábbi kolóniára jellemző baktériumközösség is átadódik az újabb kolóniába.
Ráadásul a gombakertészetet is többféle parazita támadhatja meg, sőt ugyanaz
a parazita is sokféle antibiotikum-rezisztenciával rendelkezhet. Nem biztos
tehát, hogy a korábban hatékony antibiotikum hatékony lesz az új kolóniában
is. Hogyan lehet akkor fenntartani a megfelelő antibiotikumos védekezést?
Douglas Yu kollégámmal arra
gondoltunk, hogy a kurtafarkú tintahalak által alkalmazott szűrőmechanizmushoz
hasonlóan a levélvágó hangyák is ki tudják szűrni a környezetből a számukra
kedvező baktériumokat. A jelenség nagyon leegyszerűsített modelljét dolgoztuk
ki: a gazdán két baktériumtörzs – antibiotikumot nem termelő (N) és antibiotikumtermelő
(T) – tud megtelepedni. Az antibiotikum lassítja az antibiotikumot nem
termelők növekedését, szaporodását, azonban az antibiotikum termelése költséges,
ami a termelőknek okoz szaporodási hátrányt. Megmutatható, hogy ha a gazda
kevés tápanyaggal látja el a baktériumokat, akkor mindig az N törzs fogja
uralni a baktériumtenyészetet. Azonban, ha elegendően sok tápanyagot juttat
a hangya a baktériumok számára, akkor a két törzs kezdeti arányától függően
egyensúlyban vagy a T törzs vagy az N törzs egyedei lesznek túlsúlyban
(4.
ábra). (A jelenség magyarázata durván az, hogy a tápanyagmennyiség
növelésével a termelt antibiotikum mennyisége is növekszik, annak hatása
viszont egy kritikus koncentráció fölött nagyon gyorsan nő, mert van egy
szint, ami fölött lesz igazán mérgező az antibiotikum.) Azaz, ha a hangya
a környezetből veszi fel a baktériumokat, akkor is nagy eséllyel lesznek
olyan hangyák, melyek antibiotikum-termelő baktériumokat fognak nevelni.
A szűrő nagyon egyszerű: etetni kell a baktériumokat rendesen, a többit
megoldja a baktériumok közti versengés.
 |
 |
4. ábra. A hangya által
szolgáltatott tápanyag mennyisége határozza meg, hogy az antibiotikumot
nem termelő (N) és a termelő (T) baktériumok közül melyik törzs kolonizálja
a hangya kültakaróját nagy eséllyel. A bal oldali ábra a kevés tápanyag,
a jobb oldali a sok tápanyag esete. A képen a Cyphomyrmex longiscapus levélvágó
hangya látható.
A kültakarón látható
apró fehér csomók a baktériumtelepek
Baktériumok az emberben
Talán meglepő, de durván
tízszer annyi baktériumsejttel élünk együtt, mint ahány sejt alkotja a
testünket. Ne felejtsük el: egy baktérium úgy százszor kisebb, mint egy
eukarióta sejt, így ennek a rengeteg baktériumnak a tömege csupán 1–2 kg.
Az ember emésztőrendszerében legnagyobb mennyiségben a vastagbélben fordulnak
elő, ahol szerepük igen összetett. A vastagbélben élő mikrobaközösség
(mikrobióta) segítségével emésztődik meg a táplálékkal felvett keményítő.
A vastagbél mikrobiótája, mely kb. 400–500 baktériumfajt és milliliterenként
108–1012 baktériumsejtet tartalmaz, a keményítőt
(és más poliszacharidokat) erjedés során elsősorban rövid láncú zsírsavakká
(butirát, propoinát, acetát stb.) bontja, melyek a vastagbélből felszívódnak
és megemésztődnek.
Láttuk tehát, hogy a vastagbél mikrobiótája aktív szerepet játszik a tápanyagfelvételben De van-e ezen kívül más funkciója az emésztőrendszer baktériumközösségeinek? Azt már elég régen megfigyelték, hogy erős antibiotikumos kezelés után a páciensek hajlama az emésztőrendszeren keresztül terjedő fertőzésekre ugrásszerűen megnő. Így régóta gyanítják, hogy az egészséges emésztőrendszer mikrobiótája akadályozza a paraziták elszaporodását. S valóban, az utóbbi időben egyre több kísérlet bizonyítja ezt a feltételezést. Ismereteink alapján három (egymást nem kizáró) mechanizmus segítségével védekezik a mikrobióta a kórokozók ellen: közvetlen gátlással, tápanyagelvonással és a menedékek elfoglalásával, valamint az immunrendszer aktivációjával.
Közvetlen gátlás: Ha a mikrobióta baktériumtörzsei között vannak olyanok, melyek specifikus antibiotikumokat termelnek, akkor azok gátolják az ilyen anyagokra érzékeny törzsek terjedését. (Ahogy a levélvágó hangyáknál már láttuk.)
Tápanyag elvonása és a menedékek elfoglalása: Mivel egy rendkívül fajgazdag, alkalmazkodott mikrobióta van jelen az emésztőrendszerben, az ott élő fajok nagyon hatékonyan használják fel a tápanyagokat. Emellett filmet alkotva beborítják a bélrendszer nyákos felszínét. Ez utóbbi azért fontos, mert az emésztőrendszeren keresztülhaladó táplálék és folyadék kimosná az ott élő baktériumokat, ha ez a menedékül szolgáló biofilm nem lenne. A kórokozók tehát nagyon hatékony versenytársakkal kell, hogy megküzdjenek, melyek nem sok táplálékot és menedéket hagynak a betolakodóknak.
Az immunrendszer aktivációja: Az emésztőrendszerben lévő baktériumokat az immunrendszer természetesen észleli. A megszokott mikrobióta azért is fontos, mert így az immunrendszer éber állapotban lesz, s fertőzés esetén sokkal gyorsabban tud hatékony immunválaszt elindítani.
Egy japán kutatócsoport nemrég igazolta, hogy egérben az Esherichia coli nagy eséllyel halálos fertőzést (súlyos hasmenés, vastagbélgyulladás) okozó törzse ellen egyes bifidobaktériumok hatásos védelmet nyújtanak. Ez az E. coli törzs egy speciális Shiga néven ismert toxint termel, ez okozza a gyulladást. Ráadásul a Shiga a bélfalon átjutva, a véráramba kerülve mérgezést is okoz. A bifidobaktériumok egyrészt a vastagbél immunrendszerét megfelelően stimulálják, másrészt az általuk a keményítő bontása során előállított nagymennyiségű acetát és a baktérium speciális enzimei megakadályozzák, hogy a Shiga a bélből a véráramba jusson. Továbbá a megnövekedett acetát szint olyan géneket kapcsol be a gazdában, melyeknek gyulladáscsökkentő hatásuk van. A bifidobaktériumok hatékonyságát jelzi, hogy ha az E. coli fertőzést hét nappal megelőzően ezeket a törzseket adták az egereknek szájon át, akkor szinte mind életben maradtak, ezzel szemben a bifidobaktériummal nem kezelt egerek 5–7 nap alatt elpusztulnak. Tehát a bifidobaktériumok közvetlen gátlással és az immunrendszer aktivációjával is védenek a kórokozók ellen.
Korábban már bemutattuk, hogy a levélvágó hangyák hogyan tudnak nagy eséllyel számukra kedvező baktériumokhoz hozzájutni. A gerinceseknél is felmerül ez a kérdés: Hogyan választja ki a gazda a megfelelő mikrobiótát. Az emlősöknél (s így az embernél is) tudjuk, hogy az újszülött emésztőrendszerében nincsenek mikróbák, azokat a környezetből veszi fel a születése utáni néhány napban. Az újszülött emlős születése után az anyában levő baktériumokat kapja meg. A kedvezőtlen mikrobiótájú, s így beteges anyák utódai is nagyobb eséllyel lesznek betegesek és gyengék, a megfelelő mikrobiótájúaknak viszont életképesebb utódaik lesznek. Így a gazda számára kedvezőbb mikrobióták egyre nagyobb eséllyel lesznek megtalálhatóak a populációban. Az is igaz, hogy ilyenkor alakul ki az újszülött állat immunrendszere is, mely fokozatosan „megtanulja” elfogadni az emésztőrendszerben lakó baktériumokat, s közben gyomlálja a veszélyesnek tűnő törzseket. Az emlősökre elmondottak természetesen az emberre is érvényesek. Csecsemőkön végzett vizsgálatok alapján tudjuk, hogy a stabil mikrobióta csak egyéves kor körül alakul ki.
Összegzés
A mikrobák számára a magasabb
rendű élőlények speciális stabil élőhelyet jelentenek. Ezeken az élőhelyeken
működésében és fajösszetételében is rendkívül gazdag élőközösségeket alakítanak
ki. Az eukarióták metabolizmusa nagyon konzervatív, a baktériumoké igen
sokrétű, ezért a baktériumok a stabil élőhelyért és tápanyagokért cserébe
sokszor életfontosságú szolgáltatásokkal fizetnek. Igyekeztem érzékeltetni,
hogy a gazda–mikrobióta együttélés nem valami éteri harmónia, inkább egy
jó kompromisszum a gazda és a mikrobák számára is. Mivel a gazdaszervezetek
között is szelekció van, ennek hatására olyan mechanizmusok jelentek meg,
melyek segítségével a gazda nagy eséllyel távol tudja tartani a káros baktériumokat,
miközben számára kedvezőbb baktériumközösség kialakítását segíti elő.
Bemutattam a nitrogénfixáló baktériumoknak az egész bioszféra szempontjából kulcsfontosságú szerepét, és részletesen elemeztem a rhizobium növénykapcsolatban a gazdanövény kölcsönhatás előtti és utáni válogatását, mint lehetséges szűrőmechanizmust. Azt is megemlítettem, hogy a csaló és együttműködő baktériumtörzsek közötti verseny nem szükségszerűen vezet a csalók teljes győzelméhez: ha a nyereség az együttműködők gyakoriságának telítődő függvénye, akkor a csalók és az együttműködők együtt fognak élni a gazdában. A kurtafarkú tintahal és a V. fisheri, valamint a levélvágó hangyák és a kutikulájukon élő baktériumtelepek mutualizmusát elemezve felvetettem, hogy a számára kedvező törzseket előnyben részesítő körülményeket alakít ki a gazda, kiszűrve így a „nem vágyott” betolakodókat. Hangsúlyoztam, hogy az emlősök (ember) nagyon komplex mikrobiótát tartanak fenn az emésztőrendszerükben, mely mikrobióták nemcsak a tápanyaglebontásban, hanem a kórokozók elleni védelemben is döntő szerepet játszanak.
Bizonyára érzékelte az olvasó, hogy számos kérdésre egyelőre csak tapogatózó válaszokat tudunk adni, azonban az már most is biztosan látszik, hogy az evolúciós és ökológiai szemlélet a rohamosan terjedő géntérképezési eljárásokkal megerősítve rövid időn belül forradalmi szemléletváltást fog okozni a gazda–mikrobióta kölcsönhatás megértésében. Nem kétséges, hogy ezek a felfedezések a gyógyászatra is nagy hatással lesznek. Izgatottan várjuk a fejleményeket.
A cikkben bemutatott saját
kutatásokat az OTKA K100299-es számú pályázata támogatta.
| Szószedet
Antibiotikum-rezisztencia: Olyan gének vagy géncsoportok, melyek hatására a gének hordozója védetté válik az adott antibiotikummal szemben. Baktériumfaj: Hasonló tulajdonságokkal (külső megjelenés és metabolikus) rendelkező sejtek összessége. Eukarióta: Valódi sejtmaggal, sejtszervecskékkel, sejtvázzal rendelkező sejt. Evolúciós stabilitás: A mutációval vagy génátrendeződéssel megjelenő új típusok terjedésével szembeni ellenállás. Mikrobióta: Mikroba (baktériumok, mikrogombák, vírusok) fajok populációinak együttese, melyek egyazon élőhelyen (pl. gazdában) élnek egyazon időben. Mutualizmus: Különböző fajokhoz tartozó egyedek kölcsönösen előnyös kapcsolata. Populáció: Azonos fajhoz és azonos szaporodási közösségbe tartozó egyedek összessége. Prokarióta: Sejtmaggal, sejtszervecskékkel, sejtvázzal nem rendelkező, az eukariótánál ősibb sejtek. Az eukarióták bizonyos prokarióták tartós együttéléséből jöttek létre. |
Irodalom
Archetti, M. és Scheuring
I. 2011. Coexistence of cooperation and defection in public goods games,
from the prisoner’s dilemma to the volunteer’s dilemma. Evolution 65: 1140-
Fukuda, S. és mtsai.
2011. Bifidobacteriacan protect from enterophatogenic infection through
production of acetate. Nature, 469: 543-
Kiers, E. T. és Denison
R. F. 2008. Sanctions, cooperation and the stability of plant-rhizosphere
mutualisms. Annu. Rev. Ecol. Evol. Syst. 39: 215-
McFall-Ngai, M. J. 2008.
Quick Guide: The Hawaiian Bobtail Squid. Curr. Biol. 18: R1043-.
Scheuring, I. and Yu, D.
W. 2012. How to assemble a beneficial microbiome in three easy steps. Ecology
Letters. 15:1300-
Stecher, B. és Hardt W.
2011. Mechanisms controlling pathogen colonization of the gut. Curr. Opinion
in Microbiol. 14:82
| Természet Világa, | 144. évfolyam, 2. szám,
2013. február
http//www.termvil.hu/ https://www.chemonet.hu/TermVil/ |