TRÁJER ATTILA
Ősföldtani
zátonytörténelem
Az állatok szervezetén belüli, irányított ásványképzés képessége számos, független fejlődési vonalon alakult ki a törzsfejlődés során. A biomineralizáció célja belső, illetve külső váz képzése. Amennyiben ezek a folyamatok kontroll nélkül, nem megfelelő anatómiai helyen zajlanak le az élő szervezetekben, ásványkiválások jöhetnek létre, mint amilyenek a különböző testüregekben megjelenő „kövek”. (Ezek kémiai összetétele jóval változatosabb, mint a vázak képzésében résztvevő ásványok alkotóinak száma.) Az élő szervezetek elhalása után a vázak ellenállóbb anyaguknál fogva megmaradhatnak, és fontos alkotói lehetnek a később esetleg kőzetté váló (diagenizálódó) üledékeknek. Sok esetben felismerhető az eredeti biológiai struktúra, máskor csak ún. biohermákban marad felismerhető enyhe vagy közepes fokú metamorfózist követően az eredeti szerkezet maradványa (ahogy ez történt pl. a Szendrői-hegység devon koralljaival és tengerililiom-vázaival).

1. ábra. Különböző korokban képződött sztromatolitok. Bal
felső kép: 2,2 milliárd éves sztromatolit metszete; jobb felső kép:
felső-triász korú sztromatolitos dolomit; alsó kép: devon időszakban képződött
sztromatolit (A szerző felvételei)
A
továbbiakban röviden áttekintjük a zátonyok történetét, kezdve a prekambrium
cianobaktériumok (Cyanobacteria) alkotta meszes képződményeitől a modern kori
korallalga-zátonyokig. Áttekintjük a légköri szén-dioxid szintjének a
zátonyképződésre kifejtett hatását és megismerjük az óceánok jelenkori
savasodásának előképeit a földtörténetben.
Mintegy
2,7–2,2 milliárd évvel ezelőtt a légköri szén-dioxid-koncentráció mintegy
százszorosa volt a jelenleginek. [2] A légköri szén-dioxid kivonásában
meghatározó szerepe volt a fontoszintetizáló cianobaktériumoknak. Az
asszimiláció során keletkezett szerves anyagok egy része a tengeri üledékbe
kerülve eltemetődött, szenet vonva ki a geokémiai körforgásból. Feltehető, hogy
a fotoszintézis miatt csökkenő légköri szén-dioxid-szinthez a cianobaktériumok
fotoszintetikus apparátusa folyamatosan alkalmazkodott. A légköri szén-dioxid
megkötésében azonban egy másik folyamat, a karbonát-produkció is szerepet
játszott. [3]
A
mai tengerek vize enyhén alkalikus (pH~7,5–8,4), ami azonban a Föld őskorában
nem volt így. Az archaikumra és a proterozoikum első felére jellemző igen magas
atmoszferikus szén-dioxid-koncentráció mellett a savas óceánok vizének szabad
szén-dioxid és szénsav koncentrációja a mai állapothoz képest számottevő volt.
Ennek következtében a kalcit- és aragonit-telítettségi értékek alacsonyak
lehettek, ami nem kedvezett az élő szervezetek karbonát-anyagú vázépítésének. A
cianobaktériumok fotoszintetikus szén-dioxid megkötése növelte a sejtjeik
közvetlen környezetének pH-ját, ami elősegítette lokálisan a kalcium-karbonát
biogén kiválását a baktériumtelepek mátrixában. [3] Ennek oka a szén-dioxid
vízben való oldódásának mikéntjében keresendő. A szén-dioxid disszociációja
vizes közegben a következő egyenlettel írható le:
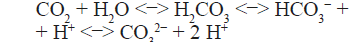
Ezek
szerint a szén-dioxid számos gáztól (pl. oxigén, nitrogén) eltérően nemcsak
fizikai, hanem kémiai úton is oldódik. Alacsony pH-n a szabad szén-dioxid és a
szénsav mennyisége magasabb lesz, ami nem kedvez a karbonátképződésnek. Ezzel
ellentétben, ha a pH alkalikus irányba változik, akkor az egyensúly az egyenlet
jobb oldala felé, a karbonátok képződésének lehetősége irányába mozdul el. Ha a
fotoszintézis szénsavat vagy szén-dioxidot távolít el, bekövetkezik a biogén
mészkiválás, mivel a vízben jól oldódó kalcium-hidrogén-karbonáttal (Ca(HCO3)2)
szemben a kalcium-karbonát (CaCO3) oldékonysága kifejezetten rossz. Ez a
folyamat ma is megfigyelhető pl. a szilvásváradi Szalajka-patak vagy a
felsőörsi Malom-patak völgyében.
A
légköri szén-dioxid koncentrációjának lassú csökkenésével a proterozoikum
folyamán az extracelluláris kalciumkarbonát-kristályok kicsapódása a
cianobaktériumok környezetében egyre könnyebben ment végbe. A karbonátok
képződése felgyorsult, amikor a légköri szén-dioxid parciális nyomása a mai
szint 33-szorosának megfelelő szintre esett le, és a kicsapódó kalcium-karbonát
mésziszap formájában kezdett felhalmozódni a tengerfenéken mintegy 1,4–1,3
milliárd évvel ezelőtt [3]. Megjegyzendő, hogy mészkövek kisebb mennyiségben
már korábban is képződtek, sőt egyes grönlandi mészkövek átkristályosodására és
márvány képződésére már 1,8 milliárd évvel ezelőtt is sor került. A
sztromatolitok által létrehozott zátonyképződmények több száz méteres
vastagságot érhettek el olykor. Az észak-amerikai Nagy-Rabszolga-tó partjait
alkotó, közel 2 milliárd éves fosszilis zátony még ma is jelentős vastagságú.
Típusos, a prekambriumi sztromatolitokra emlékeztető képződmények jelenleg pl.
a nyugat-ausztráliai Cápa-öbölben figyelhetők meg, azonban a Föld több pontján,
elsősorban túlsós vízű trópusi-szubtrópusi lagúnákban máshol is előfordulnak
sztromatolitok (a Bahamákon, a Perzsa-öbölben). Maga a Cápa-öbölben képződő
recens sztromatolitzátony is egy meleg vízű lagúnában tenyészik. A hiperszalin
vízben nem élnek meg az algákat fogyasztó állatok. Feltehetően éppen az állatok
megjelenése vetett véget a sztromatolitzátonyok korának, és alapozta meg a
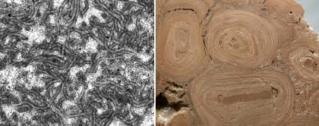
későbbi nagy fanerozoikumi zátonyképződmények létrejöttének alapját (2. ábra).

2. ábra. Egy 3,5
milliárd éves fosszilis sztromatolit Ausztráliából (North Pole; bal oldali kép)
és recens sztromatolitok a Cápa-öbölben, szintén Ausztráliában (jobb oldali
kép)
Paleozoikum
3. ábra. Girvenella fosszilis telepének szálas
szerkezetű (filamentózus) szövete elektronmikroszkóp alatt (bal oldali kép) és
a telep makroszkópos megjelenése (jobb oldali kép: onkoidok)
A
fotoszintetizáló szervezetek szén-dioxid-megkötő mechanizmusának fejlődése
700–570 millió évvel ezelőtt, a „hólabda-föld” korszakában (az elnevezés a
globális szintű eljegesedésre utal) lelassult, mivel a hideg tengervíz
elősegítette a fizikailag oldott szén-dioxid pusztán diffúzió által történő
felvételét a sejtekbe [3]. Ezzel egy időben megszűnt a meszes sztromatolit
zátonyok képződése a Földön. A nagy jégkorszak után felmelegedő bolygón
ismételten megjelentek a sztromatolitok, és nagy kiterjedésű kékalga-zátonyok
léteztek a neoproterozoikumban is. Ez a folyamat egyben elősegíthette az első
állatok megjelenését és fejlődését, valamint az állati szervezetben megvalósuló
mészvázépítés kialakulását is. Bár az első ismert állati szervezetek
testfosszíliái „csak” mintegy 610 millió évesek [5], a molekuláris óra és egyes
vitatott nyomfosszíliák alapján az állati szervezetek megjelenése 1 milliárd
évvel ezelőtt történhetett. [6] Valószínűbb, hogy inkább a fosszilizációra
alkalmas mészváz vagy meszes kitinváz aránylag hirtelen megjelenéséről van szó
a kambrium elején, ami valóban „egy időben” (ami több tíz millió éves intervallumot
is jelenthet több, mint fél milliárd év távlatából nézve), de függetlenül
jelentkezett több állatcsoport esetében is. Az ún. „kambriumi robbanás”
lényegében a mai állattörzsek hirtelennek tetsző megjelenését jelentette a
neoproterozoikum és a kambrium határán. Az első kagyló- és ízeltlábú-fosszíliák
542 millió évesek, a kambriumi robbanás „címerállatai”, a trilobiták pedig csak
21 millió évvel később, mintegy 521 millió évvel tűntek fel az élet színpadán.
Az első, állatok által létrehozott zátonyok alkotói a vitatott rendszertani
hovatartozású, 525 millió éve megjelent kora kambriumi Archaeocyathák voltak (a
kambrium első felét kb. 510 millió éve lezáró kihalást a menedéket jelentő ún.
refúgiumokban túlélték és a késő kambriumig éltek). [7] Serlegszerű vázuk
leginkább a mai mészszivacsokéval vethető össze. Érdekessége ennek a kihalt
csoportnak, hogy a ma ismert kambriumi, szilárd vázzal rendelkező szervezetek
diverzitásának több mint a felét ezek a szervezetek tették ki. [8] A
kambrium-ordovícium határán, 488 millió éve lezajlott kihalás után hosszú időn
keresztül nem épültek állatok alkotta zátonyok a világtengerekben, és helyettük
csak a kékalgák által épített thrombolit-, onkoid- és sztromatolit-zátonyok
nyújtottak élőhelyet a zátonylakó szervezeteknek. A többsejtű állatok alkotta
zátonyok újbóli globális szintű elterjedése a szilur időszak közepére tehető,
amikor a csalánozók egyes csoportjai körében olyan evolúciós folyamat ment
végbe, ami meghatározónak bizonyult a következő 450 millió év zátony-ökoszisztémáinak
szempontjából. A későbbi zátonyok felépítésében jelentős szerepet játszó
korallok már az ordovíciumban megjelentek, de jellemző zátonyalkotókká a szilur
időszakig nem váltak, köszönhetően részben az ordovícium-szilur időszak
határán, 450–440 millió évvel ezelőtt súlyos kihalási eseménynek, ami a
zátonyalkotó korallokat, pörgekarúakat, mohaállatokat és tüskésbőrűeket
különösen súlyosan érintette. Az akkori korallok nem sorolhatók be egyik ma élő
csoportba sem. Az ún. Tabulata korallok inkább telepalkotók, a Rugosa korallok
jellemzően magányos polipáriumokat építő alakok voltak.
A
szilur-devon zátonyok fontos alkotói voltak a sztromatoporoideák, amelyek
fennmaradt szerkezetük alapján a szivacsokkal mutatnak rokonságot.
Magyarországon devon időszaki korallok ismertek a Szendrői-, és ritkaságként az
Upponyi-hegységből; sztromatoporoidea-maradványok a balatonfői Polgárdi Mészkő
felső, biohermás tagozatából. A sztromatoporoideák elsősorban nem az ún.
frontzátonyt, a tengeri áramlatoknak leginkább kitett, a nyílt tengerre tekintő
zátonyrészt, hanem a zátony belsejében fekvő lagúnák felé eső zátonyterületet
népesítették be (4. ábra).

4. ábra. Szilur-devon
időszaki zátonyalkotó szervezetek maradványai. Bal felső kép: Favosites
tabulata korall metszete a szilurból; jobb felső kép: tabulata koralltelep a
devonból; bal alsó kép: erősen átkristályosodott, feltehetően
sztromatoporoidea-telep metszete, devon; jobb alsó kép: esőmarta devon zátonymészkő-feszín

5. ábra. Devon
időszaki tengeri liliomok nyéltag-maradványai és a teljes élőlény
rekonstrukciója

6. ábra. Ivanovia
filloid alga (bal oldali kép) és Caninophyllum rugosa koralltelep (jobb oldali
kép) a karbon időszakból
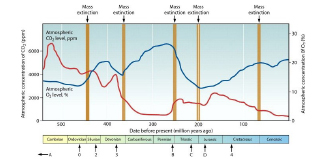
8. ábra. A légköri
oxigén- és szén-dioxid-koncentráció változása a fanerozoikum során Dorell és
Smith (2011) alapján az öt legjelentősebb kihalási esemény feltüntetésével
A
triász elején egyáltalán nem képződtek zátonyok. Ezek újbóli létrejötte a
középső-triász anisusi korszakára tehető, amikor egyes mészalgák, mohaállatok,
mészszivacsok alkottak zátonyokat elsősorban a Nyugat-Tethys
karbonátplatformjának a nyílt óceánhoz legközelebbi peremterületein. [19] E kor
emléke az ún. diploporás mészkő és dolomit, ami a hajdani, ún. dasycladacea
mészalgák csövecskéinek kioldódását követően vált lyukacsossá. A középső-triász
időszak anisusi emeletében képződött Tagyoni Formáció (Dunántúli-középhegység)
szubtidális (hullámverés alatti) fácieszében gyakoriak ezek az algák.
Physoporella, Oligoporella és Teutloporella nemzetségek fajai kerültek elő (9.
ábra).

9. ábra. Physoporella
sp. és rekonstrukciója (a kép jobb felső sarkában)
Az
egyelőre még magányos kőkorallok is csak a középső triászban tűntek fel, melyek
a mai korallzátonyok meghatározó (névadó) alkotói. Feltehetően ekkor jött létre
a csalánozók és az algák között az a szimbiotikus kapcsolat, ami a csoport
sikerének alapját jelentette és jelenti ma is. A krétában egy „helytülő”
(szesszilis), egyedülálló módon serleg vagy tülökformát felvett héjú
kagylócsoport, a Hippuritidae család tagjai alkottak zátonyokat. Telepeik,
szemben a korallok formálta képződményekkel, nem annyira vertikálisan, mint
horizontálisan voltak nagy kiterjedésűek. Olyan kagylókból származtak,
melyeknek eredetileg szimmetrikus héjai megcsavarodtak, majd az egyik héjfél
zárólemezzé, a másik egy serleg formájú képletté alakult. A héjat pórusok törték
át, amin keresztül feltehetően víz átáramoltatására is lehetőség volt. Nem
kizárt, hogy a Scleractinia korallokhoz hasonlóan szimbionta algáik is voltak.
A kréta időszak során a trópusi tengerekben visszaszorították a korallokat és
nem egy estben a zátonyok domináns alkotóivá váltak. A zátonyalkotó kagylók
kréta időszaki sikere és a korallok felett gyakorolt időleges dominanciája
mögött ismételten atmoszférikus és klimatikus okok fedezhetők fel. Egyrészt, a
kréta időszak folyamán a trópusi tengerek 6–14 °C-kal melegebbek voltak a
maiaknál, másrészt, magasabb volt a sótartalom, ami nem kedvezett a
koralloknak. [20] Ehhez jön még az, hogy a légköri szén-dioxid-koncentráció a
mai érték hatszorosa, közel 1700 ppm lehetett, jelentős savasodást okozva az
óceánokban. Az ún. rudista kagylók tündöklésének azonban vége lett, mert előbb
a kréta időszak végére megfogyatkoztak, majd a kréta/tercier határon történt
tömeges kihalás során teljesen kivesztek az élővilágból. Azóta csak az
osztrigák körében találunk kisebb zátonyépítő fajokat. A Hippuritidae kagylók
maradványai egyes hazai kréta időszaki kőzetekben gyakoriaknak mondhatók (10.
ábra).
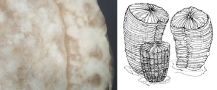
10. ábra. Hippurites
sp. metszete (bal oldali kép)
és az élő kagylók
rekonstrukciója (jobb oldali kép)
Kainozokum
Láthattuk,
hogy a múlt tükröt tart a jelen és a jövő felé. Ha tovább folytatódik az
óceánok elsavasodása, az a perm-triász határon észlelt világméretű
zátonypusztuláshoz hasonló eseményhez vezethet, ami súlyosan hatna a
világtengerek biodiverzitására. Mint láttuk, a régmúlt korokban évmilliókat,
akár 20–30 millió évet is igénybe vett a zátonyközösségek újjászerveződése, így
egy mai kihalási hullám után emberi léptékkel mérve mérhetetlen hosszú idő
telne el az új zátony-ökoszisztémák felépüléséig még egy antropogén hatásoktól
mentes világban is.

Irodalom
[2]
Rye, R., Kuo, P. H., & Holland, H. D. (1995). Atmospheric carbon dioxide
concentrations before 2.2 billion years ago. Nature, 378(6557), 603.
[3]
Riding, R. (2006). Cyanobacterial calcification, carbon dioxide concentrating
mechanisms, and Proterozoic–Cambrian changes in atmospheric
composition. Geobiology, 4(4), 299-316.
[4]
Haas, J. (2004). Magyarország geológiája. Triász (Geology of Hungary.
Triassic.
[5]
Costa, James T.; Darwin, Charles (2009). The annotated Origin: a facsimile
of the first edition of On the origin of species. Harvard University Press.
p. 308. ISBN 978-0-674-03281-1.
[6]
Campbell, Neil A.; Reece, Jane B. (2005). Biology (7th ed.). Pearson, Benjamin
Cummings. p. 526. ISBN 978-0-8053-7171-0.
[7]
Debrenne, F. (1991). Extinction of the Archaeocyatha. Historical
Biology, 5(2-4), 95-106.
[8]
Pálfy, J. (2000). Kihaltak és túlélők (Félmilliárd év nagy
fajpusztulásai)[Victims and Survivors (Mass Extinctions in the Past Half
Billion Years)].
[9]
Kershaw, S., Li, Y., Crasquin-Soleau, S., Feng, Q., Mu, X., Collin, P. Y.,
Reynolds, A., & Guo, L. (2007). Earliest Triassic microbialites in the
South China block and other areas: controls on their growth and
distribution. Facies, 53(3), 409-425.
[10]
Ezaki, Y., Liu, J., & Adachi, N. (2003). Earliest Triassic microbialite
micro-to megastructures in the Huaying area of Sichuan Province, South China:
implications for the nature of oceanic conditions after the end-Permian
extinction. Palaios, 18(4), 388-402.
[11]
Baud, A., Richoz, S., & Marcoux, J. (2005). Calcimicrobial cap rocks from
the basal Triassic units: western Taurus occurrences (SW Turkey). Comptes
Rendus Palevol, 4(6), 569-582.
[12]
Pruss, S. B., & Bottjer, D. J. (2004). Late Early Triassic microbial reefs
of the western United States: a description and model for their deposition in
the aftermath of the end-Permian mass extinction. Palaeogeography,
Palaeoclimatology, Palaeoecology, 211(1), 127-137.
[13]
Hips, K., & Haas, J. (2006). Calcimicrobial stromatolites at the
Permian–Triassic boundary in a western Tethyan section, Bükk Mountains,
Hungary. Sedimentary Geology, 185(3), 239-253.
[14]
Weidlich, O., Kiessling, W., & Flügel, E. (2003). Permian-Triassic boundary
interval as a model for forcing marine ecosystem collapse by long-term
atmospheric oxygen drop. Geology, 31(11), 961-964.
[15]
Retallack, G. J. (2001). A 300-million-year record of atmospheric carbon
dioxide from fossil plant cuticles. Nature, 411(6835), 287.
[16]
Zachos, James C., et al. „Rapid acidification of the ocean during the
Paleocene-Eocene thermal maximum.” Science 308.5728 (2005):
1611-1615.
[17]
Pandolfi, J. M., Connolly, S. R., Marshall, D. J., & Cohen, A. L. (2011).
Projecting coral reef futures under global warming and ocean
acidification. science, 333(6041), 418-422.
[18]
Kleypas, J. A., Feely, R. A., Fabry, V. J., Langdon, C., Sabine, C. L., &
Robbins, L. L. (2005, April). Impacts of ocean acidification on coral reefs and
other marine calcifiers: a guide for future research. In Report of a
workshop held (Vol. 18, p. 20).
[19]
Senowbari-Daryan, B., Zühlke, R., Bechstädt, T., & Flügel, E. (1993).
Anisian (Middle Triassic) buildups of the Northern Dolomites (Italy): the
recovery of reef communities after the Permian/Triassic crisis. Facies, 28(1),
181-256.
[20]
Johnson, C. C. (2002). The Rise and Fall of Rudist Reefs: Reefs of the dinosaur
era were dominated not by corals but by odd mollusks, which died off at the end
of the Cretaceous from causes yet to be discovered. American
scientist, 90(2), 148-153.
[21]
Báldi, T. (2003) Egy geológus barangolásai Magyarországon: Kiadó: Vince
Kiadó Kft., Budapest.
Képek
forrása
2.
netsaver.myds.me és https://www.sharkbayvisit.com.au/stromatolites.aspx
3.https://www.fossilmall.com/EDCOPE_Enterprises/stromatolite/
/stromatolites46/stromatolites-46.htm
5.
https://www.dcfossils.org
7. https://elte.pene.hu/!foldtud/2.%20felev/bevezetes_a_foldtortenetbe/
/Ftori_BSc1_11.pdf
8.
ec.asm.org
9.
https://paleopolis.rediris.es/cg/CG2013_B01/CG2013_B01_Pl_07_14.htm
| Természet Világa, | 148. évfolyam, 9. szám,
2017. szeptember http//www.termvil.hu/ |