Petesejt-citoplazma és anyai hatás
Az állatvilágban általános jelenség, hogy a petesejt, pete, vagy tojás térfogata messze meghaladja a spermiumét. Az embernél „csak” kétszázezerszer, a baromfiaknál pedig sokmilliárdszorosan nagyobb a petesejt. Vajon miért a nagy különbség, ha a női és a hímivarsejt is csak egy-egy kromoszóma-garnitúrával járul hozzá az új élőlény képződéséhez? Az általános vélekedés szerint azért, mert a női ivarsejtek citoplazmája sziket tartalmaz, olyan anyagokat, amelyek fedezik az embrió energia- és tápanyagszükségletét. A „szikhipotézis” értelme nyilvánvaló a petét (tojást) rakó élőlények életében, ahol az utódok fejlődését a pete anyagai biztosítják. De vajon miért nagyobbak az olyan élőlények petesejtjei a spermiumoknál, amelyek embriói az anyaméhen belül fejlődnek, ahol lehetőség van az embrió (magzat) folyamatos ellátására? Azért, mert a peték, a petesejtek citoplazmája nemcsak sziket, hanem olyan anyagokat is tartalmaz (mindenekelőtt mRNS- és fehérjemolekulákat), amelyek a petesejt érése során kerülnek a petesejtek, a peték citoplazmájába, és elsősorban ezek irányítják az embriógenezis kezdeti lépéseit. A jelenséget anyai hatásnaknevezik, és szerepe sokkal fontosabb életünkben, mint bárki hinné. Gondolta volna a kedves olvasó, hogy az anyai hatás a magzatvédő program alapja?
Az anyai hatás
embriológiai bizonyítékai
Régi megfigyelés, hogy sok féreg- és zsákállatfajnál a pete citoplazmáját mozaikos elrendeződében különböző színű részek alkotják (1. ábra). A citoplazmarészek az embriógenezis folyamán az embrió más-más sejtjeinek válnak a részévé, és úgy tűnik, hogy meghatározzák a sejtek sorsát. Például a Stylea partita zsákállat izmai azokból a sejtekből fejlődnek, amelyekbe a pete citoplazmájának barna része kerül, jelezve, hogy a barna citoplazma meghatározza az izomképződést. Valóban, ha egy petéből kiszívjuk a sötétbarna részt, az embriónak nem képződnek izmai. Ha a kiszívott citoplazmát egy másik pete akármely részébe injektáljuk, olyan embrió képződik, amelynek szokatlan helyen is képződnek izmai. Az itt említett egyszerű kísérletek eredményei megmutatták, hogy a pete citoplazmájának sötétbarna részében valóban vannak olyan anyagok, amelyek meghatározzák a sejtek izomtípusú fejlődését. A Stylea partita az ún. mozaiktípusú fejlődés szép példája: az embriósejtek fejlődési programját olyan molekulák határozzák meg, amelyek anyai eredetűek, és amelyek a petesejt citoplazmájában mozaikszerűen vannak elrendezve.
1. ábra. A Stylea partita zsákállat petesejtjének citoplazmáját
mozaikszerűen elrendezett, különféle színű citoplazmarészek alkotják
A muslica (Drosophila melanogaster) és a békapeték egy részében ún. csíraplazma van, abban pedig RNS- és fehérjemolekulákból álló rögöcskék (2. ábra). Azok a sejtek differenciálódnak ősivarsejtekké, amelyeknek az embriógenezis folyamán a csíraplazma a részévé válik. Az ősivarsejtekből képződnek majdan a kifejlődő élőlény ivarsejtjei. Azt, hogy valóban a csíraplazma határozza meg az ősivarsejtek képződését, K. Illmensee bizonyította az 1970-es évek elején. Illmensee vékonyka tűvel csíraplazmát szívott ki egy vadtípusú (barna testszínű) muslica petéjéből. A csíraplazmát olyan pete szokatlan részébe injektálta, amely sárga testű nősténytől származott. Mintegy két óra elteltével a sárga színű nőstény petéjében nemcsak a megszokott helyen képződtek ősivar küllemű sejtek, hanem máshol is. Azt, hogy e sejtek valóban ősivarsejtek, Illmensee a következő kísérletben bizonyította. A szokatlan helyen képződött sejteket vékony üvegtűvel összegyűjtötte, majd olyan gazdaembrió ősivarsejtjei közé ültette, amelynek szülei fekete testűek voltak. Az imágóvá fejlődött gazdát – azt eldöntendő, hogy milyen típusú ivarsejtjei képződtek – sárga partnerrel pároztatta. Ha a beültetett („sárga”) sejtek valóban ősivarsejtek és képződnek belőlük ivarsejtek, a fekete testű gazdamuslicának és sárga testű partnerének lesznek sárga utódai. (A saját „fekete” és a „sárga” ivarsejtek egyesüléséből barna utódok származnak.) Minthogy Illmensee kísérletében voltak olyan fekete gazdamuslicák, amelyektől a barnák mellett sárga utódok is származtak, nyilvánvaló, hogy a csíraplazmában valóban vannak olyan tényezők, amelyek meghatározzák az ősivarsejtek, és a majdani utódok képződését.
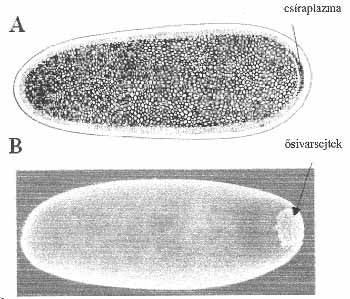
2. ábra. (A) A muslica pete hátulsó végében csíraplazma van, benne RNS- és fehérjemolekulákból álló rögöcskék. (B) Azokból a sejtekből, amelyekbe a csíraplazma kerül, ősivarsejtek képződnek
Az anyai hatást bizonyítják azok a békazigóták is, amelyek sejtmagját ha eltávolítják, egy ideig mégis úgy fejlődnek, mint azok a testvéreik, amelyeknek megvannak saját génjeik. Bár a sejtmag nélküli békaembriók fejlődése egy idő után lelassul, mielőtt elpusztulnak, már néhány száz „sejtből” állnak, és némi fantáziával ebihalaknak tűnnek. Tehát a pete citoplazmája eleve tartalmazza mindazokat az anyai eredetű anyagokat, amelyekre az embriógenezis kezdetén szükség van.
Azoknak a békaembrióknak a fejlődése, amelyek aktinomicin-D oldatban fejlődnek, megegyezik a sejtmag nélküli embriókéval. (Az aktinomicin-D olyan antibiotikum, amely megakadályozza a gének transzkripcióját, azt, hogy a génekben tárolt genetikai információ kifejeződjön.) Az aktinomicin-D-s kísérlet azt jelzi, hogy a békaembriók az egyedfejlődés kezdetén nem használják saját génjeiket, csupán az anyai eredetű anyagokra hagyatkoznak. Vajon mi a helyzet az olyan élőlényekkel, amelyek embriói az anyaméhen belül fejlődnek? Nos, aktinomicin-D-oldatban pl. az egérembriók fejlődése akkor áll meg, amikor négy sejtből állnak. Vagyis az egérembriók fejlődése során a négysejtes stádiumban van először szükségük valamely saját génjükre. Aktinomicin-D-oldatban az emberi embriók fejlődése már a kétsejtes állapotban elakad, ami csupán azt mutatja, hogy az embrió saját génjeit korán kezdi használni, de biztosan nem jelenti azt, hogy az anyai hatás szerepe a kétsejtes stádiumban véget érne.
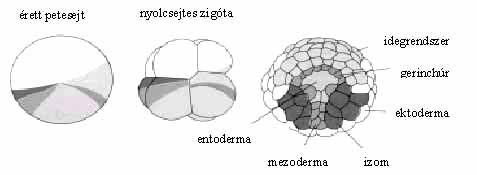
Az anyai hatást az a tény is mutatja, hogy a muslicaembriók fejlődése során anélkül történik tizenhárom osztódás és alakul ki a nagyjából hatezer sejtből álló blasztoderma (hólyagcsíra), hogy az embrió saját génjei aktiválódnának és a bennük tárolt genetikai információ kifejeződne. A különös az, hogy a blasztodermasejtek fejlődési programja már többé-kevésbe meghatározott: „tudják”, hogy utódsejtjeikből milyen szövetek, szervek fejlődnek ki (3. ábra). Nyilvánvaló, hogy a blasztodermasejtek sorsát meghatározó anyagok részei a petesejt citoplazmájának. Mik az anyai hatást közvetítő anyagok? Hol vannak a pete citoplazmájában? Hol, mikor és hogyan képződnek? Mit csinálnak és hogyan? Mi a szerepük az embriók életében? Csupa válaszra érdemes kérdés.
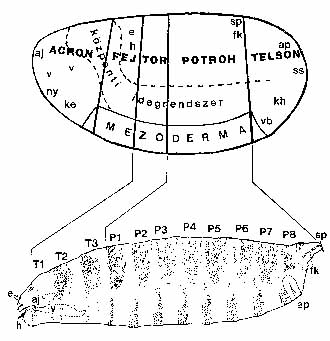
3. ábra. A muslica blasztoderma-sorstérképe azt ábrázolja, hogy a blasztodermasejtek leszármazottaiból a majdani lárva mely sejtjei képződnek. (A sorstérképet úgy készítik, hogy az embrió néhány sejtjét egy jól definiált helyen ultraibolya lézersugárral elpusztítják, majd meghatározzák, hogy a lárva testének mely sejtcsoportja hiányzik. A T és a P betűk a tor-, illetve a potrohszelvényeket jelölik. A kisbetűk a lárva néhány jellegzetes struktúráját szimbolizálják.)
Mindezek tisztázására, 1985-ben H. G. Frohnhofer vékony tűvel fiatal muslicaembriók elülső részébe szúrt, majd a kibuggyanó cseppnyi citoplazmát eltávolította (4. ábra). A „lékelt” petében olyan embrió fejlődött, amelynek hiányzott a feje. A „lékelés” eredménye meglepő, hisz a fej olyan blasztodermasejtekből származik, amelyek a sorstérképen messze vannak a pete elülső végétől, attól a helytől, ahonnan a citoplazma kifolyt. Nyilvánvaló, hogy a pete citoplazmájának elülső részében olyan anyai eredetű anyag van, amely meghatározza a fej képződését.
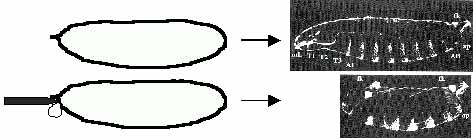
4. ábra. (A) Egy muslicapetében olyan lárva fejlődik, amelynek jellegzetes testtájai vannak. (B) Egy csepp citoplazma folyik ki abból a muslicapetéből, amelynek elülső részébe beleszúrunk. Az elején “meglékelt” petében olyan lárva fejlődik, amelynek nincs feje. Fej nélküli lárvák képződnek a bcd/bcd mutáns nőstények petéiben is, és nem képződik feje hb/hb homozigóta embrióknak sem
Az anyai hatás
genetikai bizonyítékai
Az öröklődés alapvető törvényszerűségeit először Georg Johann Mendel
ismerte fel és közölte 1865-ben. Röviden: az A/A sárga homozigóta
és az a/a zöld homozigóta borsó keresztezéséből származó
minden utód A/a heterozigóta és sárga. (A és a
ugyanannak a génnek egy-egy változata, allélja. A domináns, a
recesszív.)
Az A/a x A/a keresztezésből a második generációban
A/A, A/a és a/a
utódok származnak
1:2:1 arányban, a sárga:zöld borsók aránya pedig 3:1. A keresztezés eredménye
nem függ attól, hogy A/A (vagy a/a) nő- vagy
hímnemű: az A/A![]() x a/a
x a/a![]() és
az a/a
és
az a/a![]() x
A/A
x
A/A![]() ún.
reciprok keresztezések eredménye azonos.
ún.
reciprok keresztezések eredménye azonos.
A Mendel-szabályok 1900-ban történt újrafelfedezése után arra figyeltek
fel a genetikusok, hogy néhány tulajdonság öröklődése különleges. Egyebek
között a Limnaea peregra nevű mocsári csigafaj házának tekeredési
iránya is (5. ábra). Abban a keresztezésben, amelyben jobbra tekeredő
házú nőstény szülőket (D/D![]() )
balra tekeredő házú hímekkel (d/d
)
balra tekeredő házú hímekkel (d/d![]() )
kereszteztek, az F1 utódok (D/d) háza jobbra tekeredett,
ahogyan az Mendel első szabálya szerint várható. Nyilvánvaló, hogy D
domináns, d recesszív. Meglepő módon azonban az F1 utódok keresztezéséből
(D/d x D/d) származó minden F2 utód háza jobbra
tekeredett. Partner hiányában a Limnaea peregra hímnősként viselkedik.
A hímnős módon szaporodó F2 csigák 3/4-ének csupa jobbra tekeredő házú
utóda lett, 1/4-énél viszont minden utódnak balra tekeredett a háza. Nyilvánvaló
tehát, hogy az F2 csigák között voltak d/d-k, de valamiért
mégis jobbra tekeredett a házuk. Az is világos, hogy a csigaház öröklődése
követi a Mendel-szabályokat, csak egy generációval megkésve.
)
kereszteztek, az F1 utódok (D/d) háza jobbra tekeredett,
ahogyan az Mendel első szabálya szerint várható. Nyilvánvaló, hogy D
domináns, d recesszív. Meglepő módon azonban az F1 utódok keresztezéséből
(D/d x D/d) származó minden F2 utód háza jobbra
tekeredett. Partner hiányában a Limnaea peregra hímnősként viselkedik.
A hímnős módon szaporodó F2 csigák 3/4-ének csupa jobbra tekeredő házú
utóda lett, 1/4-énél viszont minden utódnak balra tekeredett a háza. Nyilvánvaló
tehát, hogy az F2 csigák között voltak d/d-k, de valamiért
mégis jobbra tekeredett a házuk. Az is világos, hogy a csigaház öröklődése
követi a Mendel-szabályokat, csak egy generációval megkésve.
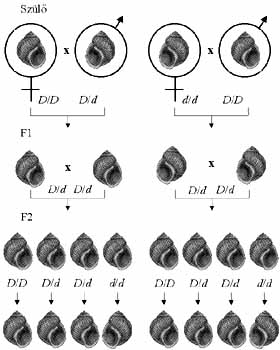
5. ábra. A Limnaea peregra csiga házának tekeredési iránya jellegzetes példa az olyan tulajdonságok öröklődésére, amelyeket anyai hatás határoz meg
Meglepő módon, a reciprok keresztezésből, amelyben balra tekeredő házú
nőstény szülőket (d/d![]() )
jobbra tekeredő házú hímekkel (D/D
)
jobbra tekeredő házú hímekkel (D/D![]() )
kereszteztek, az F1 utódok (D/d) háza balra tekeredett (5.
ábra). A különös jelenség magyarázata egyszerű: bár minden F1 csiga
D/d,
házuk tekeredési irányát nem a saját, hanem anyjuk genetikai állománya
határozza meg.
)
kereszteztek, az F1 utódok (D/d) háza balra tekeredett (5.
ábra). A különös jelenség magyarázata egyszerű: bár minden F1 csiga
D/d,
házuk tekeredési irányát nem a saját, hanem anyjuk genetikai állománya
határozza meg.
A csigaház tekeredése attól függ, hogy tesz-e vagy sem az anya a petesejtjének citoplazmájába olyan fehérjét, amely meghatározza a magorsórendszer irányát a zigóta első sejtosztódásai során. Ha tesz (a nőstény hordozza a D allélt), az utód háza jobbra tekeredik. Ha nem tesz (a nőstény d/d), az utód háza balra tekeredik. Minthogy annak a fehérjének a szintézisét, amely a magorsórendszer irányát meghatározza, az anya génjei kódolják, érthető, hogy a csigaház tekeredési irányát anyai eredetű, a pete citoplazmájában levő fehérje határozza meg.
A csigaház tekeredési irányának különleges öröklődése hívta fel először
a figyelmet az anyai hatás jelentőségére. Lássunk egy további példát. Csupa
fertilis, szaporodni képes utódok származnak azokból a keresztezésekből,
amelyekben vadtípusú (+/+) nőstény muslicákat kereszteznek gs/gs
hímekkel. A reciprok keresztezésből (gs/gs![]() x +/+
x +/+![]() ) származó utódok viszont
annak ellenére sterilek, hogy hordozzák az ép (+) allélt. A gs/gs
nőstényeknek bár vannak utódaik, nincsenek unokáik. (Innen a gs
mutáció neve: grandchildless.) Minthogy a gs/gs
nőstények
petéiből hiányzik a csíraplazma, érthető, hogy bár a gs mutáció
recesszív, a gs/+ utódoknak mégsem képződnek ivarsejtjeik, sterilek.
Ma már tudjuk, hogy az ép (+) gén terméke a csíraplazma egyik alkotója.
) származó utódok viszont
annak ellenére sterilek, hogy hordozzák az ép (+) allélt. A gs/gs
nőstényeknek bár vannak utódaik, nincsenek unokáik. (Innen a gs
mutáció neve: grandchildless.) Minthogy a gs/gs
nőstények
petéiből hiányzik a csíraplazma, érthető, hogy bár a gs mutáció
recesszív, a gs/+ utódoknak mégsem képződnek ivarsejtjeik, sterilek.
Ma már tudjuk, hogy az ép (+) gén terméke a csíraplazma egyik alkotója.
A genetikai boncolás
és zigóta génjei
Az említett példák azt mutatják, hogy az anyai hatás létező jelenség; a petesejtek, a peték (tojások) citoplazmájában vannak olyan anyai eredetű anyagok, amelyek meghatározzák az embrió sejtjeinek, és végeredményben az utódok sorsát. Vajon valóban van a muslicapete citoplazmájában a „feji részt” meghatározó anyag? Hogyan lehetne azt az anyagot, illetve szerepét megismerni? A megoldást az ún. genetikai boncolás módszere adja a következő gondolatmenet alapján. Ha van olyan gén, amelynek terméke része a pete citoplazmájának, és szükséges a fejképződéshez, akkor mutációval megszüntethető a gén funkciója. Arra számíthatunk, hogy a mutációra homozigóta (mutáns) muslicanősténynek bár lesznek petéi, de ha megtermékenyülnek is, bennük fejetlen embriók képződnek. A fejetlen embriók nem tudnak kikelni a petékből, elpusztulnak. Tehát a mutáns nőstények végül is sterilek. Ha tehát azt a gént akarjuk azonosítani, amelynek terméke a fej képződéséhez szükséges, először a nőstények sterilitását okozó, ún. nőstény-steril mutációkat kell létrehozni, majd közülük kell kiválogatni azokat, amelyeknél az embrióknak nem képződik feje. A mutációk lényegében olyan eszközök, amelyekkel ép géneket azonosíthatunk. A mutáns fenotípus (itt a fejtelenség) alapján lehet következtetni az ép gén funkciójára. Ahhoz, hogy megismerhessük az azonosított ép gén molekuláris funkcióját, klónozni kell az ép gént.
A „vadászat” a fej képződéséhez szükséges génekre az 1970-es évek végén kezdődött C.Nüsslein-Volhard és E. Wieschaus laboratóriumában, Heidelbergben. Először nőstény-steril mutációkat indukáltak, majd kiválogatták közülük azokat, amelyek petéiben fej nélküli embriók képződtek. A nőstény-steril mutációk három olyan gént azonosítottak, amelyek funkciója a fej képződéséhez szükséges: a bicoid- (bcd), az exuperentia- (exu), és a swallow- (swa) géneket. A bcd/bcd mutáns nőstények petéiben csupa fej nélküli embrió képződik. Olyanok, mint amilyenek az elejükön meglékelt petékben fejlődtek. Kiderült, hogy a három gén közül a bicoid-gén szerepe a legfontosabb. A bcd mutációból kiindulva Nüsslein-Volhard és munkatársai (Tübingenben, az 1980-as évek második felében) előbb meghatározták az ép bicoidgén helyét a muslicagenomban, majd klónozták a gént azért, hogy megismerjék molekuláris szerepét a fejképződésében.
A kutatók a bicoid mRNS-t a pete citoplazmájának elülső részében találták meg. A bicoid mRNS-molekulák a sejtvázhoz rögzülnek, pontosan oda, ahonnan a lékelés során a petékből kifolyik egy kevéske citoplazma. A bicoid mRNS-ről röviddel azelőtt képződnek a bicoid fehérjemolekulák, mielőtt a blasztoderma kialakulna, akkor, amikor a sejtek fejlődési programja meghatározódik. A bicoid fehérjekoncentrációja fokozatosan csökken a pete elülső csúcsától a hátulsó vég felé, egy ún. gradiens mentén. C. Nüsslein-Volhard és munkatársai azt is megmutatták, hogy a bicoid-fehérje adott koncentrációja az embriósejt meghatározott típusú fejlődési programját határozza meg. Azt, hogy a bicoid fehérje valóban morfogénként hat, az mutathatja, hogy a sejtmagvakban halmozódik fel, és feltételezhető, hogy úgy szabályozza a sejtek fejlődési programját, hogy az embrió génjei közül némelyeket be-, másokat kikapcsol.
Ha a bicoid fehérje valóban bekapcsolja a zigóta valamely génjét, mutációval megszüntethető a bekapcsolt gén funkciója. A bicoid-szabályozott gén funkciójának hiányában arra számíthatunk, hogy a mutáns embriónak nem képződik feje, és elpusztul. A fej nélküliség azonban nem az anya, hanem az embrió genotípusától függ. A Drosophila génjei között mindössze egy van, az ún. hunchback (hb) gén, amelynek mutációi megfeleltek az említett elvárásnak: a hb/hb mutáns embrióknak nem képződik fejük és elpusztulnak. A hunchback gén funkciójának kiesése miatt az embrió testének jellegzetes része – mint egy rés – hiányzik; a hb gén az ún. „gap” (rés) gének jellegzetes példája.
Nem meglepő, hogy a hb gén azokban a sejtekben fejeződik ki (vagyis képződik bennük Hunchback fehérje), amelyekből a fej fejlődik. Minthogy a bcd/bcd mutáns nőstények embrióiban nem expresszálódik a hb gén, nyilvánvaló, hogy a bicoid fehérje valóban bekapcsolja a hb gént. A Krüppel (Kr) rés-gén az embrióknak azokban a sejtjeiben expresszálódik, amelyek hb gént expresszálók mögött következnek. Minthogy a Kr gén a bcd/bcd mutáns nőstények embrióiban az elülső sejtekben is expresszálódik, nyilvánvaló, hogy a Bicoid fehérje gátolja a Kr gén expresszióját az embrió elülső sejtjeiben. A Bicoid fehérje valójában olyan transzkripciós faktor, amely a hb és a Kr gének promóterében a DNS jellegzetes bázispársorrendű részéhez kötődik, és ha alkalmas koncentrációban van jelen, a hb-gént be-, a Kr gént kikapcsolja. Tehát a bicoid mRNS és a Bicoid fehérje közreműködésével a nőstény határozza meg – anyai hatás révén – embriói sejtjeinek sorsát.
Anyai hatás az emlősökben
Az anyai hatás szerepéről az emlősökben ma még nagyon keveset tudunk,
de, hogy szerepet játszik benne, két megfigyeléssel illusztrálom. Amíg
egy lókanca és egy szamárcsődör hibridje, az ún. lóöszvér a ló anyára,
a szamárkanca és a lócsődör hibridje, az ún. szamáröszvér, a szamár anyára
hasonlít. Bár a kétféle öszvér génjeinek egyik fele ló, a másik fele szamár
eredetű, küllemük annyira különbözik, hogy sok nyelvben külön szavakat
is használnak a kétféle öszvérre (6. ábra). Angolban a lóöszvér
neve mule, a szamár-öszvéré hinny. Különös, de ma sem tudjuk,
hogy a kétféle öszvérbe honnan, hogyan és mikor származik a „lóság” és
a „szamárság”, mikor és hogyan határozza meg a kanca azt, hogy az öszvér
utód rá hasonlítson.
 |
 |
6. ábra. A ló kancától származó öszvér a ló anyára (balra), a szamár kancától származó öszvér a szamár anyára hasonlít testmérteiben (jobbra)
Régi megfigyelés, hogy a csikók testmérete az anyákéra emlékeztet. A nagy testű (ún. sodrott, mint a muraközi) kancák és a szokásos méretű csődörök keresztezéséből származó csikók csaknem kétszer akkora súllyal születnek, mint a reciprok keresztezésből származó csikók. Bár mindkét csikóféleség génjeinek fele sodrott, a másik fele normál lótól származik, a testtömeg mégis eltérő, és az anyáéra emlékeztet. Ugyan a különbség eredete nem ismert, a háttérben minden bizonnyal az anyai hatás áll.
A közelmúltban derült fény arra, hogy az emberi petesejtek citoplazmája is tartalmaz olyan molekulákat, amelyek a korai embriógenezist szabályozzák. Nyilvánvaló, hogy elpusztulnak az embriók, ha az anyai eredetű tényezők valamelyike hiányzik, vagy a szokottnál csekélyebb mennyiségben van a petesejt citoplazmájában. Két fő oka lehet annak, hogy egy emberi petesejt nincs tökéletesen feltöltve az anyai eredetű tényezőkkel.
A genetikai ok azt jelenti, hogy egy asszony homozigóta egy olyan m mutációra, amely miatt petesejtjeinek citoplazmájából hiányzik egyfajta anyai eredetű géntermék. A genetikai ok is kétféle lehet. (1) A funkcióját vesztett génnek az egyetlen szerepe az, hogy a petesejt-citoplazma valamely komponensét kódolja. (Mint pl. a muslica bicoid, vagy grandchildless génjei.) Nyilvánvaló, hogy az m/m mutáns asszonyok testi funkciói épek, sőt képződnek petesejtjeik, amelyek meg is termékenyülnek. De bárki is legyen az m/m asszony partnere, zigótáiban el sem kezdődik az embriógenezis, vagy ha mégis, hamarosan leáll, az embriók elpusztulnak tehát, az asszony meddő. (2) Vannak olyan – mondjuk m’ – mutációk is, amelyekben a génfunkció csak részlegesen vész el, az m’/m’ asszonyok mégis meddők. Oka az, hogy az ún. ivarsejtvonal-sejtek „genetikai igénybevétele” sokkal nagyobb, mint a testi sejteké. Bár az m’/m’ asszonyok testi sejtjei épnek, egészségesnek tűnhetnek, petesejtjeik funkciója csökkent, és alkalmatlan arra, hogy belőlük új élet kezdődjön. Az m’ típusú mutációk jelentőségét mi sem mutatja jobban, mint az, hogy a nőstény-steril mutációk 90 (!) százaléka olyan m’ típusú mutáns allél, amely a funkció részleges vesztésével jár. Ha ugyanabban a génben a funkció teljes vesztésével járó ún. zigotikus letális mutációk keletkeznek, azok az élettel összeegyeztethetetlenek. A „genetikai igénybevétel” két dolgot jelent. Egyrészt azt, hogy vannak gének, amelyek termékeinek intenzívebb a szerepe a nőstény ivarsejtekben, mint a testi sejtek bármelyikében. Másrészt azt, hogy a petesejtek citoplazmájában sokkal több gén terméke van jelen és tölt be funkciót, mint valamely testi sejtben. Amíg például a muslicapeték citoplazmája a gének legalább háromnegyedének (!) termékét tartalmazza, a testi sejtekre jellemző arány mindössze egyharmad. Ismerve az anyai hatás jelenségét és értve a gének szerepét a petesejtek fogamzásra történő felkészítésében, nem véletlen, hogy becslések szerint a meddő asszonyok 8-10 százaléka genetikai okok miatt meddő.
Az is bizonyosra vehető, hogy vannak környezeti tényezők, amelyek gátolják az anyai hatás anyagainak bekerülését és elrendeződését a petesejtek citoplazmájában. A környezeti tényezők közül a dohányfüst, valamint a szerves oldószerek azok, amelyek hatása a legdrámaibb. Sőt, a petesejtek tökéletes felkészítése nemcsak környezeti ártalmak „eredményeként” maradhat el, hanem pl. vitaminhiány miatt is. A tökéletlenül felkészített petesejtekből fejlődő magzatok között gyakori a nyitott gerinccsatorna, valamint más fejlődési rendellenesség. Az ún. magzatvédő programok célja a petesejtek tökéletes „felkészítése” a fogamzásra. Ma már tudott (sajnos, nem elég széles körben), hogy a folsav (egy koenzim, a B2 vitaminkomplex fontos komponense) a petesejtek „feltöltődésének” legfontosabb tényezője. Nem véletlen, hogy a folsav a magzatvédő vitaminkomplex egyik komponense, és az sem, hogy azoknak, akik részt vettek a magzatvédő programban, nem született nyitott gerinccsatornájú gyermekük, és utódaik között a fejlődési rendellenességek is ritkák voltak. A folsav fontosságát felismerve, ma már pl. az Egyesült Államokban a pékárukat (nálunk a táltos kenyeret) folsavval dúsítják. Az ideális állapotot természetesen az jelentené, ha tisztában lennénk azzal, hogy az anyai hatás, az utódok vállalása roppant fontos dolog. Akkor lehet esélyünk arra, hogy ép és egészséges gyermekünk születik, ha asszonyaink petesejtjeit tökéletesen feltöltötték mindazokkal az anyagokkal, amelyek az embriógenezis korai eseményeit irányítják. A folyamat mintegy három hónapig tart. Mindenkinek tudni illene, hogy a foganás előtti hónapokban tartózkodni kell a környezeti ártalmaktól, és vitamindús ételekkel kell táplálkozni. Végtelenül szomorú, hogy sok asszony csak akkor hagyja abba a dohányzást, amikor rádöbben arra, hogy egy új életet hordoz méhében, amikor már jószerivel eldőlt magzata sorsa, amikor már „lekéste” nemcsak a petesejt felkészítését, hanem magzata fejlődésének kezdeti és legfontosabb szakaszait is. Az írás összeállítója sehogy sem érti azokat a dohányos asszonyokat, akik gyermeket vállalnak, de dohányzásukkal eleve hátrányos helyzetbe hozzák saját utódaikat, akikért élnek, dolgoznak.
Sokan és sokszor megkérdezték már a „muslicásoktól”, mi értelme a „muslicakutatásoknak”,
mit tanultunk belőlük. A válasz egyszerű. A sejtek, a kromoszómák, a DNS
és a fehérjemolekulák alapvető szerkezete és funkciója nem különbözik az
emberétől. (Hinné a kedves olvasó, hogy a muslica és az ember importin-b
molekuláiban ugyanazokon a helyeken az esetek 61 százalékán ugyanazok az
aminosavak vannak, 80 százalékban pedig hasonló természetűek?) A muslicán
gyűjtött molekuláris szintű törvényszerűségek nagyjából igazak az emberre
is. A muslicákból kiindulva olyan alapvető jelenségek molekuláris mechanizmusát
ismertük meg, mint a testszerveződés szabályozása, a belső óra és a napi
ritmus, a rövid távú memória, a daganatképződés, vagy – amint talán a fentiekből
kitűnik – a meddőség genetikai alapjai.
| Természet Világa, | 133. évfolyam, 12. szám, 2002. december
https://www.chemonet.hu/TermVil/ https://www.kfki.hu/chemonet/TermVil/ |
Vissza a tartalomjegyzékhez