
E rovat írásaival olvasóink, tanáraink
átfogóbb tájékozódását igyekszünk elôsegíteni.
| MOLEKULÁRIS BIOLÓGIA | |
|
E rovat írásaival olvasóink, tanáraink átfogóbb tájékozódását igyekszünk elôsegíteni. |
CSERMELY PÉTER
Hogyan viselhetõ el az élet 95 oC-on?
A vulkanikus hõforrások, gejzírek, a tenger alatti kitörések hidrotermális áramlatai számos, eddig kevéssé ismert élõlénynek, a hipertermofil* baktériumok változatainak adnak otthont. Hogy hogyan válik az élet elviselhetõvé ilyen szélsõséges körülmények között, arra a túlélést segítõ mechanizmusok vizsgálata adhat választ. A kutatási eredmények nemcsak a földi élet keletkezésének megértéséhez, hanem saját szervezetünk jobb megismeréséhez is közelebb vihetnek minket, sõt megteremthetik a lehetõségét számos ipari, biotechnológiai alkalmazásnak és felkészíthetik az emberiséget egy esetleges idegen életforma megértésére is.
A Yellowstone Nemzeti Park gejzírjeiben, vagy a tenger mélyén, számos vidáman szaporodó baktérium, ún. hipertermofil baktérium él, amelyek szemlátomást igen jól alkalmazkodtak a megváltozott körülményekhez [1].
| 1. táblázat. A magasabb hõmérsékleten
élõ egysejtûek csoportjai |
|||
| Növekedési hõmérséklet (oC) | |||
| Min. | Opt. | Max. | |
| Termofilek | >30 | >50 | >60 |
| Extrém termofilek | >40 | >65 | >70 |
| Hipertermofilek | >70 | >90 | <115 |
A víz forrásához közeli illetve a mélytengeri magas nyomásviszonyok közepette akár a 100 fokot jóval meghaladó hõmérsékleten (1. táblázat) a hipertermofil baktériumok sejtjeinek mûködéséhez, szervezõdéséhez és szaporodásához nélkülözhetetlen makromolekulák szinte mindegyikét védelmezni kell. Ilyen magas hõmérsékleten legtöbbször nem az okozza a fõ problémát – mint az emberi szervezet sejtjeiben –, hogy “hogyan tudom egy kívánt reakció sebességét megnövelni”, hanem éppen ennek a fordítottja, nevezetesen az, hogy “hogyan lehet a nem kívánt reakciók sebességét csökkenteni”. Cikkemben néhány olyan tanulságos védekezési módot kívánok bemutatni, amellyel az extrém magas hõmérsékleten élõ baktériumok védelmezik a fehérjéiket, RNS, DNS molekuláikat, illetve a sejtet határoló membránt. Kitérek néhány más extrém körülmény (túlzott hideg stb.) kiváltotta alkalmazkodásra is, és példákat fogok hozni a szélsõséges körülményekhez való adaptáció megismerésébõl fakadó ipari és tudományos felhasználási lehetõségekre.
Fehérjék a víz forráspontjához közeli hõmérsékleten
A víz forrpontjához közeli, illetve néhány esetben az ezt meghaladó hõmérsékleten élõ hipertermofil baktériumok fehérjéi igen erõs kémiai bontóhatásnak, leginkább hidrolízisnek, valamint a szabályos struktúrák lebontására törekvõ “macerálóhatásnak” vannak kitéve.
|
2. táblázat. A fehérjeszerkezetet stabilizáló
mechanizmusok |
||
| Változás | Szerkezet- stabilizálás |
Kötés- elrejtés |
| pótlólagos kötések prolingazdag régiók ionpárok hidrofób kötések keresztkötések |
+ + + + |
|
| kompaktabb szerkezet tömör fehérjebelsõ hidrofób felszínek hiánya “loop”-ok hiánya alfa-hélixek stabilizálása |
+ + + + |
+ + + + |
| komplexek makromolekulákkal fehérje DNS RNS glikoziláció |
+ + + + |
+ + + + |
| komplexek “mikromolekulákkal” K+ (akár 2,3 M-ra emelt koncentrációban) Ca2+, Mg2+ -ionok ciklikus-1,2-difoszfo-glicerát di-myo-inozitol-1,1’-foszfát |
+ + + + |
+ + |
(A másodjára említett “macerálás” tudományosan fogalmazva így hangzik: a fehérjeszerkezet szétbomlásakor növekszik az entrópia, a hõmérséklet növelésével egyre jelentõsebb lesz az ebbõl fakadó szabadenergia csökken, így további bomlások várhatóak.) A kémiai bontóhatás ellen a fehérje leginkább a sérülékeny kötések mellõzésével, vagy elrejtésével védekezhet. A mellõzés szép példája a hidrolízisre hajlamos glutamin aminosav viszonylag kis mennyisége a hipertermofil A. aolicus baktérium összes fehérjéjében [2]. Más hipertermofilek enzimeiben az ugyancsak hidrolizálható aszparagin és az oxidábilis cisztein hiánya is gyakran kimutatható. Az elrejtés lehetséges módozataira a fehérjestabilizálás módjait felsoroló 2. táblázat mutat be néhány példát [3–5].
A hipertermofilekre jellemzõ fehérjék szerkezetének stabilitását változatos módon lehet biztosítani. Például: pótlólagos kötések beiktatásával (prolingazdag régiók, ionpárok, keresztkötések stb.), a szerkezet “lazaságainak”, “lötyögéseinek” minimalizálásával (még szorosabb illeszkedés a fehérje belsejében, még kevesebb hidrofób fehérjefelszín, a fehérje felszínén lévõ laza, hajtûszerû struktúrák, ún. “loop”-ok eltüntetése stb.), vagy más molekulákkal képzett komplexek külsõ merevítõ hatásával. Pótlólagos kötések például a hidrofób kölcsönhatások számának növekedésével jöhetnek létre a fehérjékben, ha a hõmérséklet emelkedik (kb. 70 oC-ig). A fehérjeszerkezet kompaktabbá válik, ha a hõstabil fehérjék felépítésében fontos szerepet betöltõ alfa-hélixek módosulnak. Ilyenkor a hõstabil alfa-hélixek két végén a hélixet lezáró “sapka” még a szokásosnál is több extra hidrogénhíddal csökkenti az addigi szabályos, helikális struktúra törésébõl fakadó sérülékenységet. Az alfa-hélixek egyenetlen töltéseloszlását pedig a pozitív résztöltéssel bíró N-terminális részeiken felszaporodó negatív töltésû aminosav-oldalláncok kompenzálják a hipertermofil fehérjék esetén [3–6].
A fokozott stabilitásnak azonban ára van. A merev struktúra ugyanis nem lesz annyira alkalmas a gyors és szabályozott katalízisre, mint a hajlékony, flexibilis [7]. Ez a “kár” azonban csak látszólagos. Egyrészt azért, mert a magas hõmérséklet a szerkezeti merevséget részben ellensúlyozza és a kémiai reakciók sebességét is jelentõsen megnöveli, másrészt mert a szabályozásban épp azok a magasabb rendû fehérje-fehérje, fehérje-DNS, illetve fehérje-RNS komplexek segítenek, amelyek a stabilizálásnak is egyik igen fontos eszközei. (Ennek a sejtszintû szabályozásnak, a “channeling”-nek részletes kifejtését l. a A sejtszerkezet sajátosságai c. részben.)
A fehérjék és a fehérjealapú enzimek molekuláris evolúciója valószínûleg éppen fordított utat járt be, mint az elõzõekben részletezett “emberi” enzimek —> hipertermofil enzimek átmenet. A kezdeti, igen kicsi és ezáltal igen merev és stabil fehérjestruktúrákból a késõbbiek során fejlõdhetett ki a mai enzimek nagyobb, flexibilisebb és ezáltal specifikusabb, aktívabb, valamint sokkal bonyolultabb kölcsönhatásokra és szabályozhatóságra képes családja [7]. Ennek a változásnak valószínûleg fontos mozgatórugója volt a Föld fokozatos hûlésével a merev, stabilizált fehérjestruktúrákat kikövetelõ evolúciós nyomás csökkenése is.
A DNS és RNS védelme magas hõmérsékleten
A nukleinsavak védelmének egyik legkézenfekvõbb eszköze a guanin-citozin párok arányának növelése. Ez a nukleotidpár ugyanis a másik lehetséges variációval, az adenin-timin (illetve RNS-ben adenin-uracil) párral ellentétben nem két, hanem három hidrogénhíddal létesít kötést, amely növeli a nukleinsav két szálának összetapadását. Érdekes, hogy várakozásunkkal ellentétben, csak a magas hõmérsékleten élõ baktériumok stabil RNS-eiben (transzfer-RNS-ben, riboszómális-RNS-ben stb.) találunk az átlagosnál magasabb arányú guanin-citozin párokat, a teljes genom DNS-ben azonban nem [2,3]. Ha ugyanis a DNS-ben 100%-nyi lenne a guanin-citozin pár, ennek a nagyon stabil DNS-nek az információhordozó képessége nulla lenne. A hipertermofil baktériumoknak ezért az adaptáció során a DNS/RNS stabilizálásának más útjait is ki kellett „találniuk”.
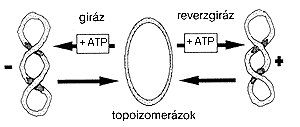 |
| 1. ábra. A DNS pozitív és negatív szupertekercsei, a reverzgiráz enzim mûködésének vázlatos mechanizmusa. ATP segítségével a giráz enzim a DNS-molekula negatív, míg a magasabb hõmérsékleten élõ organizmusokra specifikus reverzgiráz pozitív szupertekercseket hoz létre. Az ATP-vel vagy anélkül mûködõ topoizomeráz enzimek akár a pozitív, akár a negatív szuper- tekercsekkel rendelkezõ DNS-t relaxált formába alakítják vissza |
A DNS védelmének lehetséges mechanizmusa. A DNS szerkezetének védelmében eddig két fõ mechanizmust sikerült megtalálni. A stabilizálás talán legfontosabb módja az ún. „reverzgiráz” enzim mûködéséhez kötõdik, amely csak a magas hõmérsékleten élõ baktériumokban fordul elõ. A reverzgiráz mûködésének alapelvét az 1. ábra szemlélteti. A reverzgiráz a kettõsszálú DNS molekulában pozitív szupertekercseket hoz létre és ezáltal a magas hõmérsékletû élõlények DNS-ét stabilizálja. A DNS két szálának összekapcsolása (az elõbbi mechanizmustól függetlenül) szintén a DNS kettõs-láncának stabilizálódásához vezet. Ha például egy DNS-kötõ fehérje (pl. RNS polimeráz) egymáshoz rögzíti a kör alakú DNS molekula két szálának egy-egy pontját, ez már önmagában is a szálak szétválási hõmérsékletének (a DNS olvadáspontjának) akár több tíz fokos emelkedését okozhatja [3].
A DNS stabilizálásának másik fõ eszköze az az eukariota (és ezen belül: emberi) szervezetek által tökéletesített eljárás, amelyben a DNS szerkezetét a különbözõ fehérjék kötésével stabilizálják. A stabilizáció oka és célja a két szervezet esetén más és más. Amíg a hipertermofilek a magas hõmérséklet lazító hatása ellen védekeznek így, addig az eukariota sejtek a prokariota sejtekéhez képest rendkívül megnövekedett méretû (és viszonylag kis helyre, a sejtmagba összezsúfolt) DNS-üket igyekeznek ezzel a módszerrel (az így kialakuló nukleoszómák segítségével) jobban bepakolni, illetve az átíródását még szabályozhatóbbá tenni.
A legtöbb hipertermofil DNS-ének szerkezetében további stabilizációt lehetett elérni azzal, hogy a sejt a “felesleges” részeket, azaz az egyes fehérjéket kódoló gének második, harmadik és magasabb számú kópiáit, valamint a hosszú génközötti DNS-szakaszokat sorra elhagyta az evolúció során (illetve azok ki sem fejlõdtek) [3]. Száz fok körüli hõmérsékleten már a DNS kémiai módosítását (és/vagy tördelését) okozó reakciókra is fokozottan ügyelni kell. A hipertermofil baktériumok egy részében magas (akár 2,3 mólos, azaz a mi sejtjeinkben uralkodó érték több mint hússzorosát elérõ) KCl-koncentrációt, illetve olyan kétértékû kationokat (Mg2+, Zn2+) figyeltek meg, amelyek stabilizálják a DNS-t, ill. meggátolják a degradálódást is. A hipertermofilek azonban mindemellett valószínûleg hatékony DNS-javító mechanizmusokat is kifejlesztettek.
Az RNS-eket stabilizáló mechanizmusok. A sejt életében hosszabb ideig részt vevõ RNS-eket (transzfer- és riboszómális-RNS-t) a már említett guanin-citozin párokban való feldúsulás mellett (amely riboszómális RNS esetén akár a várt 50 % helyett a 70 %-ot is elérheti) a különbözõ poszttranszlációs módosítások is védik a degradáció, illetve a konformációvesztés ellen. A leggyakoribb módosítások: a különbözõ nukleotidok acetilációja és a metilezés. Ezek egyrészt stabilizálják az RNS szerkezetét azáltal, hogy kevesebb forgási szabadságot hagynak az egyes nukleotidokban elhelyezkedõ kémiai kötések körül, másrészt csökkentik elérhetõségüket, illetve megváltoztatják elektronellátottságukat, amelyek védik a különbözõ kötéseket a degradációtól. A nukleotidok módosítása mellett a kétértékû kationok (leginkább a Mg2+ -ionok) kötõdése is az RNS molekulák számottevõ stabilizálódásához vezet. A szerkezeti merevség ugyan a fehérjeenzimekhez hasonlóan az enzimaktivitással bíró RNS-ek (ribozimek) katalitikus tulajdonságainak csökkenésével jár, de ez a kémiai reakciókat amúgy is felgyorsító magashõmérsékletû környezetben valószínûleg a “kisebbik rossz”-nak bizonyult [3].
A sejtfal és a sejtmembrán hõrezisztenciája
 |
| 2. ábra. A Hyperthermus butylicus baktérium sejtfalának elektron- mikroszkópos vizsgálatával kapott elektrondenzitás-térkép. (Adaptálva a [3] irodalmi hivatkozásból.) |
A hipertermofilek sejtfalának különleges vonása. A baktériumsejtet határoló réteg az emberi sejtekre is jellemzõ sejtmembrán mellett legtöbbször egy további külsõ védõréteggel, sejtfallal is rendelkezik. A sejtfal a hipertermofil baktériumok esetén is a sejtmembránhoz kötött fehérjék és poliszacharidok komplexe. A fehérjealkotókban gyakran találunk olyan peptidkötéseket, amelyek az alfa-aminosavak oldalláncairól indulnak tovább. Ez a szokványos peptidbontó hatásoknak ellenálló kötés tovább fokozza a baktérium ellenállóságát. A sejtfal eddig sajnos még kevéssé felderített a legtöbb hipertermofil organizmus esetén, de azt már tudjuk, hogy struktúrái igen szabályos, ismétlõdõ alakzatokká állnak össze (2. ábra) [3].
A sejtmembrán adaptációja a magas hõmérséklethez. Sejtfal, illetve sejtfalszerû struktúra azonban nem található meg minden hipertermofil bakériumnál. Ezért az egysejtûek védelmében különösen fontos szerep jut a sejtet határoló sejtmembrán fokozott ellenállóképességének. A baktériumok membránjának ionok számára való átjárhatatlansága nemcsak a környezetüktõl való elszigeteltség biztosítása miatt fontos, hanem azért is, mert a sejtmembrán két oldala közötti ionkoncentráció-különbségek biztosítják azt az energiát, amellyel a baktériumok (az emberi sejtek mitokondriumaihoz hasonlóan) adenozin-trifoszfátot, ATP-t tudnak termelni.
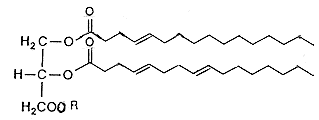
Az emberi sejtek lipidjei |
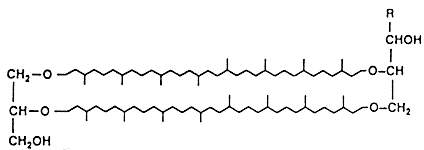 |
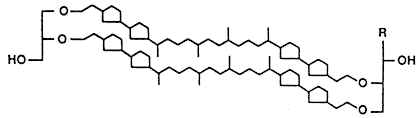
A hipertermofil sejtek lipidjei |
| 3. ábra. Az emberi sejtek membránjára
jellemzõ lipidek összehasonlítása a hipertermofil baktériumokban megtalálható lipidekkel |
Az ellenállóképesség fokozásának egyik legfontosabb eszköze az, hogy a sejtmembrán lipidjei döntõen nem észter, hanem éterkötéseket tartalmaznak. Az éterkötések ugyanis ellenállóak, nem tesz kárt bennük sem a magas hõmérséklet, sem a vulkanikus környezetben oly gyakori savas pH-n az észterkötések gyorsan lebontó hidrolízis (3. ábra). A lipidek felépítésében részt vevõ szénhidrogén-láncokban szinte soha nem találunk (a saját sejtjeinket felépítõ lipidekben oly gyakori és oly kívánatos) kettõskötéseket, ezáltal a telítetlen kettõskötések káros, a magas hõmérsékleten gyorsan végbemenõ oxidációjára sem kerülhet sor. A lipidek sajátos felépítése a “kérgüket” alkotó glicerinmolekuláknak olyan egyedi, feszített alkatot kölcsönöz, amelyet a más élõlények által kiválasztott támadó foszfolipáz enzimek nem ismernek fel. Ezáltal a speciális szerkezet révén az extrém körülmények között élõ baktériumok egy további evolúciós elõnyre tesznek szert.
Magas hõmérsékleten azonban nemcsak az egyes lipidmolekulák kémiai stabilitására kell gondot fordítani, hanem arra is, hogy az egymás mellé kerülõ lipidek összessége, a membrán maga is stabil legyen. Olyan változtatásokat kell tehát kifejleszteni, amelyek a hõmérséklet növelésével az “elfolyósodó” membránt merevebbé, az “olvasztásnak” ellenállóbbá teszik. Emiatt az éterkötésekben részt vevõ glicerinmolekulákból igen gyakran jut a lipidrészletek mindkét végére egy-egy és az így kialakuló tetraéter-vegyületek még nagyobb ellenállóságot kölcsönöznek a membránszerkezetnek. A telítetlen kettõskötések hiánya is a membránszerkezet merevítéséhez vezet. Elõfordul, hogy a szénhidrogén-láncokba ciklopentán gyûrûk ékelõdnek be, amelyek tovább csökkentik a lehetséges rotációk számát és ezáltal merevítik a létrejövõ membránszerkezetet. (3. ábra). A lipidek glicerinmolekuláihoz gyakorta kapcsolódnak cukorkomponensek és foszfát-, valamint szulfátcsoportok, hogy az extrém savas közeg fokozott polaritásához a kellõ átmenetet megteremtsék [3].
Amint azt már korábban említettem, a hipertermofilek életkörülményeire jellemzõ igen magas hõmérsékleten nem a reakciók felgyorsítása, hanem a nem kívánt reakciók megelõzése jelenti a legnagyobb gondot. Emiatt a szervezetünk sejtjeiben megszokott anyagcsereutak egymás után következõ enzimeinek sokkal szorosabban kell egymáshoz kapcsolódniuk, mint az a jelenleg szokványos földi hõmérsékleten élõ sejtek esetén megszokott. Az összetartozó folyamatokat katalizáló enzimek speciális csatolását “channeling”-nek nevezik, amelyet botcsinálta nyelvújítóként a “csõpostázás” elnevezéssel fogok illetni. A továbbiakban hangsúlyozni kell, hogy a folyamat irányított jellegû és azt is, hogy a csõpostázott anyagok a célbajuttatás során a külvilágtól elzártak. A csõpostázással meg lehet akadályozni, hogy egy reakcióláncba tartozó köztitermék “elkóboroljon” és valamely versengõ reakcióban felhasználódjon, vagy aspecifikus bomlási folyamatok áldozata legyen. Külön mechanizmusokkal kell védeni azokat a molekulákat, amelyek igen sok reakcióban vesznek részt, de egyszersmint igen reakcióképesek is. Ilyen molekula pl. az adenozin-trifoszfát az (ATP), amelynek a féléletideje védelem nélkül csupán 1 másodperc lenne a hipertermofilek szokásos hõmérsékletén [8]. A csõposta által nyújtott védelem mellett a hipertermofilek azzal is védekeznek az ATP gyors bomlása ellen, hogy az ATP-t számos reakcióban, így pl. a glikolízisben a kevésbé bomlékony, de szintén nagy energiájú adenozin-difoszfáttal (ADP), illetve pirofoszfáttal helyettesítik.
A csõpostarendszer mûködtetésére általában (így az emberre jellemzõ sejtekben is), különbözõ enzimkomplexek szolgálnak. Ezen felül azonban a hipertermofil sejteknek egy további mechanizmussal, a szokásosnál fejlettebb sejten belüli hálózattal is rendelkeznie kellene. Ugyanakkor ez a citoplazmatikus hálózat nem lehet rendkívül merev, hiszen akkor a sejt életében bekövetkezõ változásokat nehezen tudná követni. Ez lehet a magyarázata annak, hogy mindezidáig a hipertermofil sejtek belsõ szerkezetének vizsgálata kevés közvetlen eredménnyel járt. Jonathan Trent és munkatársai feltételezése szerint a hipertermofilek ezen hálózatát azok a nagy mennyiségben jelenlévõ fehérjék alkothatják, amelyek jelenlegi tudásunk szerint leginkább a formálódó vagy károsodott fehérjék helyretekerésében vesznek részt. Ezeket a fehérjéket molekuláris chaperonoknak vagy magyar elnevezéssel dajkafehérjéknek nevezzük. [9]
Alkalmazkodás a hideghez és más extrém körülményekhez
A hidegadaptáció legfontosabb kérdése, hogy hogyan tudja a sejt mozgásban tartani a mûködéséhez szükséges legfontosabb mechanizmusokat az egyre jobban szilárduló környezetben. Ahhoz, hogy a sejt reakcióit katalizáló enzimek a kémiai mozgásokat befagyasztó, hidegebb körülmények között is mûködõképesek maradjanak, a szokásosnál hajlékonyabbaknak, flexibilisebbnek kell lenniük. Ezt az élõlény a szerkezetet merevítõ prolin-aminosavak elhagyásával, és több flexibilis, hurokszerû struktúra (ún. “loop”) beiktatásával éri el. A sejt membránjai telítetlen zsírsavakban igen gazdaggá válnak, ami fluiditásukat nagymértékben növeli.
A halofil (magas sókoncentrációt szeretõ) baktériumoknál az egyik legnagyobb probléma az élethez nélkülözhetetlen víz megszerzése. A baktériumokat körülvevõ sós víz Na+- és Cl--ionjai ugyanis hidrátburkukkal a környezet víztartalmának jelentõs részét megkötik. Így nem csoda, hogy a halofil baktériumok sejtmembránjában a többszörös negatív töltéssel bíró lipidek igen felszaporodtak. A töltött lipidek a magas iontartalmú környezethez való fokozatosabb átmenetet teremtik meg és az ellenionként odavonzott Na+-ionok óriási hidrátburka révén vízhez juttatják a “szabad” vízben szegény környezetben élõ baktériumokat. Valószínûleg ugyanez, a víz “csapdába csalása” lehet az oka annak is, hogy e baktériumok fehérjéiben is igen sok a töltéssel rendelkezõ aminosavoldallánc. A többletként megjelenõ töltött oldalláncok általában a “szokásos” ionkoncentráció mellett kialakuló fehérjemaghoz csatlakozó “loop”-okon helyezkednek el. A magas nyomás elsõsorban a különbözõ fehérje-fehérje, illetve fehérje-RNS, fehérje-DNS komplexek disszociációját okozhatja [8]. Sajnos a magas nyomáshoz, a szerves oldószerekhez és más extrém körülményekhez történõ adaptáció molekuláris mechanizmusai még nem tisztultak le kellõképpen.
A természetes adaptív mechanizmusok felhasználása a gyakorlati életben
Az ipari termelésben felhasznált enzimek összértéke már 1996-ban is meghaladta a 2,5 milliárd dollárt [1]. Ezen ipari enzimek döntõ többsége extrém körülmények között is stabil. Termofil enzimeket találunk szinte minden mosóporban. A mai “kõmosott” farmerek követ sose láttak, sajátosan kopott megjelenésüket a termofil “celluláz 103” nevû enzimnek köszönhetik. A vizeket szennyezõ anyagoktól való megszabadulás az extrém körülmények között is aktív enzimek nélkül ma már ugyancsak elképzelhetetlen [10]. Igen sok ipari folyamat (a keményítõ hidrolízisétõl, a metán vagy másnéven “biogáz” termelésen át, az alacsony fémtartalmú kõzetekbõl történõ fémkioldásig) használ extrém körülmények között élõ baktériumokat. A DNS darabok nagy hatékonyságú felszaporítására használt polimeráz láncreakció (PCR) is egy termofil DNS-függõ DNS-polimerázt, legtöbbször a Thermus aquaticus baktérium ún. “Taq polimerázát” használja fel a DNS lemásolására az egyes DNS-megkettõzési szakaszokban (4. ábra). A PCR reakció a biotechnológiai munkálatok alaplépéseként ma már a személyazonosításnak és a klinikai diagnosztikának hazánkban is kiterjedten alkalmazott eszköze. A hipertermofil baktériumok sejtfalának ellenálló, szabályos mintázatai (2. ábra) mint szerkezeti anyagok, illetve mint molekuláris méretû szûrõk is alkalmazást nyerhetnek [11,12].
|
Polimeráz láncreakció (PCR) |
||
| Reakcióelegy: Ismeretlen DNS (kevés) DNS polimeráz (hipertermofil, kevés) oligonukleotid polimer (kétféle, sok) dATP, dCTP, dGTP, dTTP (sok) |
||
| 1. ciklus: | ||
| Ismeretlen DNS DNS polimeráz |
||
| + melegítés 95 oC-ra a DNS két láncának szétválasztására, majd hûtés |
||
| 2. ciklus: | ||
| másolt DNS 2. sz. primer |
ismeretlen DNS 1. sz. primer |
|
| + melegítés 95 oC-ra a DNS két láncának szétválasztására, majd hûtés |
||
| Termék: százezertôl akár százmillióig terjedô 1. sz. és 2. sz. primer közötti DNS-darb, valamint jóval kevesebb ezeken túlnyúló DNS |
||
| 4. ábra. A hipertermofil DNS-polimerázt
alkalmazó polimeráz láncreakció (PCR) vázlatos leírása |
||
Az alkalmazkodási mechanizmusok haszna a földi és a földön kívüli élet megismerésében
Az említett köznapi hasznosulási formák mellett azonban az extrém körülmények között élõ baktériumok által kifejlesztett túlélési mechanizmusok megértése közelebb vihet minket számos nagy jelentõségû kérdés megválaszolásához is. Az extremofilek tanulmányozása hozzásegít a földi élet kialakulásának jobb megértéséhez, hiszen több milliárd évvel ezelõtt a Földre jóval nagyobb mértékben volt jellemzõ a mai szemmel nézve “extrém” körülményeket jelentõ lokális környezet. A Mars jégsapkáján, a Jupiter víztartalmú holdján, az Európán esetlegesen elõforduló életjelenségek vagy a földönkívüli élet más lehetséges megnyilvánulásai is minden bizonnyal a földi élet extrém körülmények között kifejlõdött változataihoz fognak a leginkább hasonlítani. Így nem árt, ha szellemünket és fantáziánkat a környezetünkben található “legvadabb” példák tanulmányozásával is edzzük az elképzelhetetlen elképzeléséhez. Végezetül az extremofilek legtöbb faja az archebaktériumok országába tartozik. Az eukarioták kialakulásának legújabb elméletei szerint az archebaktériumok nem is olyan archaikusak, mint ahogy azt az elsõ névadáskor gondolták, hanem igen sok szempontból a saját sejtjeink közeli rokonai. Így az extrém körülmények között megjelenõ tulajdonságaik számos esetben olyan mechanizmusok aktiválódását jelenthetik, amelyek bennünk, emberekben is létezhetnek, de esetleg ma alárendelt, másodlagos szerepben. Ezek a másodlagos mûködési formák készen állhatnak arra, hogy nagyobb természeti változások esetén (pl. ha a Földünk rombolása katasztrófához vezet) aktiválódjanak, és újabb esélyt adjanak a túlélésre. Mindez természetesen csak feltételezés (bizonyítására, vagy elvetésére az emberi genom teljes megismerése után, kb. tíz év múlva kerülhet sor), de ha igaz is lenne, akkor sem adhat semmilyen felmentést maradék józan eszünk Föld-védõ kontrollja alól.
* Hipertermofil = magas hõmérsékletet különösen tûrõ (fil = barát kedvelõ; thermo = meleg; hiper = különösen kiemelkedõ); (görög).
Ajánlott irodalom
[1] Madigan, M.T. és Marrs, B.I. (1997) Extremophiles. Scientific
American, 66-71.
[2] Deckert, G. és mtsai. (1998) The complete genome of the
hypertermophilic bacterium Aquifex aeolicus. Nature 392, 353-358.
[3] Kates, M., Kushner, D.J. és Matheson, A.T. (1993) The biochemistry
of archaea. New Comprehensive Biochemistry vol. 26. Elsevier Science (hazai
fellelhetõség pl.: BME Közp. Könyvtár, katalógusszám:
353.221)
[4] Fágáin, C.O. (1997) Stabilizing protein function.
Springer Verlag
[5] Daniel, R.M., Dines, M. és Petach, H.H. (1996) The denaturation
and degradation of stable enzymes at high temperatures. Biochem. J. 317,
1-11.
[6] Jaenicke, R. (1996) Stability and folding of ultrastable proteins:
eye lens crystallins and enzymes from thermophiles. FASEB J. 10, 84-92.
[7] Csermely, P. (1997) Proteins, RNA-s, chaperones and enzyme evolution:
a folding perspective. Trends Biochem. Sci. 22, 147-149.
[8] Jaenicke, R. és Závodszky, P. (1990) Proteins under extreme
physical conditions. FEBS Lett. 268, 344-349.
[9] Trent, J. D. és mtsai. (1997) Chaperonin filaments: The
archaeal cytoskeleton? Proc. Natl. Acad. Sci. USA 94: 5383-5388.
[10] Pennisi, E. (1997) In industry, extremophiles begin to make their
mark. Science 276, 705-706.
[11] Cowan, D.A. (1992) Biotechnology of the Archaea. Trends in Biotechnology
10, 315-323.
[12] Coolbear, T., Daniel, R.M. és Morgan, H.W. (1992) The enzymes
from extreme thermophiles: bacterial sources, thermostabilities and industrial
relevance. Adv. Biochem. Eng. Biotechnol. 45, 57-98.
| Természet Világa, | 129. évf. 6. sz. 1998. július, 298–301. o. https://www.kfki.hu/chemonet/TermVil/ https://www.ch.bme.hu/chemonet/TermVil/ |