
| IMMUNOLÓGIA
|
|
GERGELY JÁNOS
Antigénfelismerés és jelátvitel
Hogyan szabályozza a
sejtek és a receptorok “beszélgetése”
az immunfolyamatokat?
Elsõ rész
Szervezetünket a kórokozóktól (patogénektõl) immunrendszerünk védi meg, amelyet felismerõ, információkat továbbító és a folyamatot kiváltó kórokozót elpusztítani képes végrehajtó (effektor) rendszerként szokunk jellemezni. Ennél tágabb értelmû megfogalmazásban azt is mondhatjuk: az immunrendszer legfontosabb feladata az, hogy különbséget tegyen a saját, a megváltozott felépítésû saját, illetve a nem saját struktúrák között. Amíg a saját struktúrákkal szemben a rendszer toleráns, addig a megváltozott szerkezetû saját struktúrákat, például a rákos sejteket, illetve a nem saját struktúrákat, mint amilyenek a baktériumok és paraziták, elpusztítja. Mindkét meghatározás arra utal, hogy az immunrendszer felismer, információkat továbbít és effektor funkciókat vált ki.
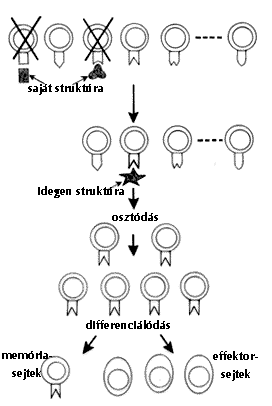 |
| 1. ábra. Immunsejtek klonális osztódása, effektor- és memóriasejtek kialakulása |
A felismerés aktív folyamat. Olyan kölcsönhatásokat jelent, amelyek során a saját vagy nem saját struktúrák felismerésére specializálódott immunsejtek aktiválódnak. Ez azt jelenti, hogy a sejt információt kap egy adott struktúráról és ezt tovább is adja olyan sejtek egész sorának, amelyek ugyanazon struktúra felismerésére specializálódtak. Ezt követõen sejtkölcsönhatások alakulnak ki, amelyek eredményeképpen az eseménysorozatba bekapcsolódó sejtek jellegzetes változásokon esnek át (1. ábra).
Ezek a változások a következõk: (1) az osztódás során felismerõ sajátságukat (specificitásukat) megõrzõ sejtek tömege (klónja) jön létre, (2) amelyek azonos felismerõ sajátsággal rendelkezõ effektor (végrehajtó) sejtekké differenciálódnak, (3) ezek egy része azonban (azonos felismerõ sajátságukat ugyancsak megõrzõ) hosszú életû, ún. memóriasejtekké alakul.
A felvázolt eseménysorozat végén tehát a folyamatot kiváltó struktúrát (antigént) felismerõ effektorsejt klón alakul ki, melynek sejtjei vagy közvetlenül képesek a patogén (nem saját, megváltozott saját) elpusztítására, vagy az antigént fajlagosan felismerõ fehérjét (ellenanyagot) termelnek és az ellenanyaggal kapcsolódó antigén a falósejtek vagy a nagy pusztítóerõvel rendelkezõ komplementrendszer áldozataivá válnak.
Az elõzõ esetben sejtközvetített (celluláris), az utóbbiban pedig ellenanyag közvetített (humorális) immunválaszról beszélünk.
Az immunválasz következménye tehát a “betolakodók” elpusztítása. Ha az immunrendszer jól mûködik, ez a pusztító folyamat célzott (fajlagos), tehát közvetlenül vagy közvetve csak az ellen az antigén ellen irányul, amely a folyamatot kiváltotta. (Tehát, ha a “betolakodó” idegen, például egy adott baktérium, az effektor mechanizmusok csak ez ellen irányulnak és az aktivált immunrendszer sejtjei és/vagy ellenanyagai csak a folyamatot megindító baktériumot pusztítják el.) Az immunrendszer azonban a “sajátot” felismerõ sejtekkel is rendelkezik: tehát alkalmas a szervezet saját építõelemeinek elpusztítására is. Joggal emlegetik tehát az immunrendszert “kétélû kard”-ként! Mûködése azonban sokrétûen szabályozott, ami több oldalról is biztosítja, hogy normális körülmények között a saját anyagokkal szemben kialakult tolerancia folyamatos és tartós legyen, hogy a pusztító folyamatot csak a szervezet épségét veszélyeztetõ struktúrák válthassák ki.
Az elmúlt három évtized kutatásai sok tekintetben tisztázták az immunrendszer mûködésének és szabályozásának molekuláris alapjait. Ennek részletes ismertetése azonban még egy tankönyv kereteit is meghaladná. Írásunkban ezért csak néhány lényeges molekuláris mechanizmust vázolunk fel, amelyek bepillantást engednek a felismerés és információtovábbítás folyamatába, valamint az immunsejt-aktiválás és az aktiváció-gátlás harmóniáját segítõ eseményekbe. Ezek során megismerheti az olvasó, hogyan “beszélgetnek” egymással az immunrendszer sejtjei, mi ennek a jelentõsége, hogyan terjed ezek eredményeként az antigénre vonatkozó információ sejtrõl sejtre, miként fognak össze a “beszélgetõ partnerek” a folyamatot kiváltó patogén fajlagos elpusztítására, valamint azt, hogyan sajátítják el egyes patogének (pl. vírusok) az immunrendszer “nyelvét”, és hogyan alkalmazzák az elsajátított mechanizmusokat saját fennmaradásuk biztosítására.
A jelátvitel
 |
| 2. ábra. Egy membránreceptor szerkezeti felépítése |
A sejt membránjához kötött felismerõ molekula, a receptor általában három részbõl épül fel: a sejten kívüli rész hordozza a kötõhelyet (ez reagál a ligandummal), a rövid, plazmamembránon áthaladó (transzmembrán) szakasz teszi lehetõvé például a receptorhoz kapcsolódó (asszociációs) molekulákkal való kölcsönhatást; a citoplazmába benyúló (citoplazmatikus) “farok” rész pedig olyan csoportokat tartalmazhat, amelyek alkalmasak a jelátviteli folyamat kiváltására (2. ábra).
A sejtek citoplazmájában egymással kapcsolódni tudó jelátvivõ molekulák sokasága található, amelyek kaszkádszerû reakciója juttatja el a membrán felõl érkezõ stimulusokat a sejtmagba. Így különbözõ aktivációs utak alakulhatnak ki, amelyek egymást kiegészítve és segítve a sejtmagba juttatják a sejtre jellemzõ, a funkciók aktivációjához (vagy gátlásához) szükséges molekulák képzõdését serkentõ jeleket. Az aktivációs folyamatok kiváltásában a protein tirozin kinázok (PTK), az aktivációs utak gátlásában pedig a foszfatázok játszanak kulcsszerepet. Az aktivációs jelek kiváltása rendszerint egy jelátvivõ molekula tirozinjának foszforilációjával veszi kezdetét. Ennek az az elõfeltétele, hogy a receptor citoplazmatikus része rendelkezzék olyan, tirozint tartalmazó molekulaszakasszal, amelynek segítségével a jelátvivõ molekulák foszforilációhoz kötött láncreakciója megindulhat.
A sejtaktiváció “lelke”: az ITAM motívum
A membránreceptorok citoplazmában elhelyezkedõ jelátvivõ szakasza jellegzetes motívumot tartalmaz, amely kulcsszerepet játszik a jelátvitel megindításában, illetve folyamatában. A fehérvérsejtek (limfociták) közé tartozó B-sejtek antigénreceptorának (BCR) jelátvivõ mechanizmusát tanulmányozva egy német immunológus a BCR-komplex járulékos láncainak citoplazmatikus szakaszában találta ezt a jellegzetes szakaszt, amelyet róla Reth-motívumnak neveztek el. Késõbb más immunreceptorok sejten belüli részén is megtalálták ezt a motívumot, amelynek szerepét a jelátvitel kérdéseivel foglalkozó kongresszuson, Kecskeméten sikerült kemény viták között végletesen tisztázni. Itt nyerte el az azóta “nagy karriert befutott” motívum az ITAM elnevezést (Immunoreceptor Tyrosine-based Activation Motif, azaz immunreceptorok tirozin alapú aktivációs motívuma).
Az ITAM általában 26 aminosavat tartalmaz (D/E – X7 – D/E – X2 – Y – X – X – L/I – X7 – Y – X – X – L/I), amelyben egymástól 6-8 aminosavnyi távolságban két, tirozin tartalmú szakasz (Y-X-X-L/I) található, ebben az “X” bármilyen aminosav lehet. A receptor-ligandum kapcsolódást követõen a tirozinok foszforilálódnak. Így a mindkét tirozinon foszforilált ITAM a jelátvivõ molekulák ún. SH2 (src-homológ) jelû doménjével kerül kölcsönhatásba, ami biokémiai események egész sorát indítja el a sejtekben. A protein tirozin kinázok mellett egyes foszfatázok is jellegzetes résztvevõi a receptorokon át kiváltott sokrétû eseménysorozatnak, amelyek folyamán fehérje–fehérje kölcsönhatások jönnek létre, jelátvivõ molekulák szerinje/treoninja foszforilálódik, másodlagos hírvivõ molekulák képzõdése révén a jelátvivõ utak aktiválódnak, végül pedig transzkripciós faktorok képzõdnek, amelyek a sejtmagba jutva specifikus gének átíródását váltják ki. Ezáltal a sejtmagban olyan folyamatok indulnak el, amelyek eredménye a sejt aktiválódása vagy éppen ellenkezõleg, anergiája (mûködésképtelensége), esetleg apoptotikus pusztulása (programozott sejthalála) lehet (3. ábra).
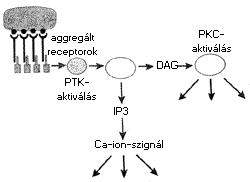
3. ábra. A jelátvitel útja a membránreceptortól
a sejtmagig
(PLC = foszfolipáz-C, PKC = szerin/treonin kináz, DAG = diacil
glicerol,
IP = inozitol foszfát (másodlagos hírvivô molekulák)
Az ITAM-függõ sejtaktiváció “gátja”:
az ITIM
Ez a szójátéknak is beillõ alcím egy “motívumpárost” jellemez, amelyet a közelmúltban egy régóta ismert folyamat tanulmányozása során ismertünk meg és amelynek magyar vonatkozása is van. Nemcsak azért, mert jó néhány közleménnyel mi is hozzájárultunk a probléma megfejtéséhez, hanem azért is, mert két évvel az ITAM kecskeméti karakterizálása után a Tihanyban rendezett nemzetközi, jelátvitellel foglalkozó kongresszuson született meg az ITIM elnevezés és a motívum jellemzése.
A B-sejtek aktivációjának (Fcg-receptorok [FcgR] útján történõ) gátlását tanulmányozva derült fény arra, hogy ennek a gátló jeleket közvetítõ koreceptornak a citoplazmatikus szakaszán egy, az ITAM-hoz sok tekintetben hasonló motívum található. Ez az ITIM-nek (Immunreceptor Tyrosine based Inhibition Motif) elnevezett motívum akkor lép mûködésbe, ha a koreceptor (FcgRIIb) és az aktivációt megindító B-sejt–receptor (BCR) között kapcsolat alakul ki. Az ITIM 13 aminosavból áll (A – E – N – T – I – T – Y – S – L – L – M – H – P), amelyen belül két aminosavval elválasztott tirozint és leucint tartalmazó résztõl függ az aktivációs folyamat gátlása. A gátlás elõfeltétele a motívumban található tirozin foszforilációja. Az elmúlt három évben kiderült az is, hogy az említett koreceptor (FcgRIIb) mellett egy sor más, gátló funkciót közvetítõ receptor is tartalmaz ITIM-et, amelyekre szintén jellemzõ, hogy:
– az ITAM-mal rendelkezõ receptorokkal történõ
kapcsolódás után gátolják az ITAM-indukált
aktivációs folyamatokat;
– a foszforilált ITIM SH2-domént tartalmazó
foszfatázokkal kerül kölcsönhatásba (ellentétben
az ITAM-mal, amelyek SH2 tartalmú kinázokkal kapcsolódnak);
– a gátló hatás annak tulajdonítható,
hogy az ITIM foszfatázokat visz az ITAM-aktivált jelátvivõ
molekulák közelébe és defoszforiláció
útján gátolja a tirozin foszforiláció-függõ
aktivációs folyamatokat.
Az antigénfelismerõ struktúrák
Tágabb értelemben antigénnek nevezünk minden olyan struktúrát (sejtet, molekulát), amelyet az immunrendszer antigénfelismerõ receptorokkal rendelkezõ T- és B-limfocitái felismernek. A limfociták membránján található antigénfelismerõ receptorok egy adott antigénnel fajlagos kapcsolatba lépve aktivációs vagy gátló jeleket továbbítanak a sejt felé.
Ahhoz, hogy a limfociták aktiválódjanak, legalább két jel szükséges (4. ábra). Az elsõ szignált az antigénreceptor és az antigén kölcsönhatása jelenti, ami a sejtet aktivációra kész állapotba juttatja. Ahhoz azonban, hogy a limfocita osztódjék és effektorsejtekké differenciálódhasson, további jel(ek)re van szükség, amely(ek) ún. koreceptorok útján jut(nak) el sejthez. A koreceptorok más sejtekkel történõ kapcsolódást, más sejtek termékeivel (pl. citokinekkel) való reakciót tesznek lehetõvé és az antigénreceptor útján felvett “fajlagos” elsõ jel a koreceptor(ok) útján kapott információkkal (második jel) együtt indítják el a sejtet az aktiváció vagy ellenkezõleg, a gátlás útján.
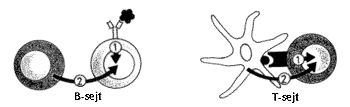
4. ábra. A limfocita-aktiváció két szignált igényel. B-sejt: az 1.szignál az antigéntôl, a 2. szignál egy T sejttôl; T sejt: az 1. szignál az antigéntôl, a 2. szignál az antigént prezentáló sejttôl származik.
Egy adott limfocita egy adott antigén felismerésére specializálódott receptorral rendelkezik. Az immunválasz feltétele tehát az, hogy az antigén a limfociták sokaságából kiválassza (szelektálja) azt a sejtet, amely a megfelelõ felismerõ receptort hordozza membránján. Ez a szelekciós folyamat általában a perifériás nyirokszervekben (nyirokcsomó, lép) zajlik le. Ha a megfelelõ antigén-receptor párok egymásra találtak, a limfociták aktiválódása következtében megindul az adott specifitású sejtek osztódása, effektorsejtekké történõ átalakulása és végül egy olyan sejtpopuláció (klón) alakul ki, amelynek minden egyes sejtje az adott antigén felismerésére alkalmas receptort hordozza membránján (1. ábra).
Háromféle antigénfelismerõ molekuláról
tudunk eddig, amelyek ugyan egymással szerkezetileg rokonok, de
mûködésük tekintetében jelentõs eltéréseket
mutatnak. Ezek a (1) B-sejt receptorok (BCR) és a velük
szerkezetileg azonos szolubilis
B-sejttermékek, az ellenanyag-molekulák; (2) a fõhisztokompatibilitási
génkomplex (MHC) termékei, amelyek minden magvas sejt
membránján megtalálhatók; (3) és a T-sejt
receptorok (TCR).
Az antigénfelismerõ molekulák szerkezeti rokonsága abból adódik, hogy valamennyien egy molekulacsaládba, az immunglobulin (Ig) szuperfamíliába tartoznak. E molekulacsalád (az Ig) szerkezetének kutatása mintegy négy évtizedre nyúlik vissza. Az Ig-ok, ezek a négy polipeptidláncból álló, heterodimer egységekbõl felépülõ, jellegzetes doménszerkezetet mutató, ellenanyag-aktivitással rendelkezõ globulinok voltak az elsõ olyan “felismerõ” molekulák, amelyek szerkezetét, polimorfizmusuk szerkezeti és genetikai hátterét, a szerkezet és funkció összefüggéseit szinte minden tekintetben sikerült tisztázni. Az utóbbi években az is világossá vált, hogy a sejtfelszíni vagy az oldott molekulák egész sora jelentõs szerkezeti homológiát mutat az Ig-okkal, továbbá, hogy ezek többsége receptorként és/vagy ligandumként felismerést, sejtkölcsönhatásokat közvetít. Ezzel együtt információtovábbító funkciót töltenek be. (Az Ig-ok szerkezetével a Természet Világa 1988. 2. számában Gergely János: Orvosi Nobel-díj, 1987 címû írása foglalkozott.)
A B-sejt receptorkomplex
Ehrlich, a századforduló zseniális immunológusa feltételezte, hogy az antigént felismerõ molekulák és a sejtek membránján található antigénreceptor szerkezetileg azonosak. Ehrlich elképzelése igaznak bizonyult, mert – mintegy fél évszázaddal késõbb – kiderült, hogy a B-limfociták által termelt ellenanyag-molekula sejtmembránhoz kötött változata tölti be az antigénfelismerõ receptor funkcióját ezeken a sejteken.
Az Ig-molekula N-terminális doménje variábilis (V) szerkezetû, így minden egyes B-sejt klón sejtjeinek membránján az antigénreceptor ún. V-doménje is eltérõ szerkezeti felépítést mutat. Mivel a molekulának ez a része tartalmazza az antigén egy meghatározott elemének (antigén determináns, epitóp) befogadására alkalmas “zseb”-et, érthetõ, hogy az egyes B-sejt klónok más és más epitóp felismerésére “specializálódtak”. A B-sejtek csontvelõi õssejtekbõl származnak, érett limfocitákká történõ fejlõdésük a csontvelõben zajlik. A sejtek egyedfejlõdése során az Ig-molekula egyes doménjeit kódoló gének véletlenszerû kapcsolódása, átrendezõdése eredményezi a sejtenként eltérõ szerkezetû V-domének, és ezen belül az adott sejtre jellemzõ antigénkötõ hely kialakulását. (A témával a Természet Világa 1992. 9. számában Uher Ferenc: Védelmi rendszer a szervezetben címû írásában foglalkoztunk részletesen.) A csontvelõben az érési folyamat mellett jellegzetes szelekciós eseménysorozat is lejátszódik, amelynek során a saját struktúrákat felismerõ B-sejtek zöme elpusztul vagy reakcióképtelen állapotba kerül (negatív szelekció). A sejtek sokaságának ez az (egész élet folyamán tartó) érése és szelekciója egy olyan B-limfocita klónkészlet kialakulását eredményezi, amely alkalmas a 109-11 nagyságrendûnek becsült antigéntömeg felismerésére.
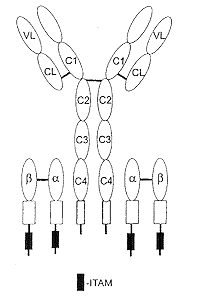 |
| 5. ábra. A BCR-komplex szerkezete |
A sejtmembránon megjelenõ receptor kettõs funkciót tölt be: egyrészt felismeri és megköti az antigént, amely több receptort képes a sejt felszínén összekapcsolni, másrészt az így keresztbekötött receptorok aktivációs vagy gátló jeleket váltanak ki a sejtben.
Amint erre az elõbbiekben utaltunk, az antigént a B-sejt membránjához kötött és ezért membrán-Ig-nak (mIg) is nevezett ellenanyag-molekula ismeri fel. Hamarosan azonban kétségek vetõdtek fel, hogy ez az mIg vajon alkalmas-e arra, hogy aktivációs jeleket továbbítson a B-sejt felé, tekintve, hogy az érett sejtek membránján antigénreceptorként legkorábban megjelenõ mIgM citoplazmatikus szakasza mindössze három aminosavat tartalmaz és ezek között nincs tirozin. Az mIgM molekula tehát jelátviteli folyamatok kiváltására alkalmatlan! Két jelentõs megfigyelés hívta fel a figyelmet arra, hogy az antigénfelismerõ hellyel rendelkezõ mIg ezen hiányossága ellenére mégis be tudja tölteni a receptorfunkciót. Kimutatták ugyanis, hogy a négy polipeptid láncból felépülõ mIg-hoz két heterodimer molekula (Ig-a-Ig-b) kapcsolódik, továbbá, hogy az Ig-a és Ig-b citoplazmatikus részében foszforilációra alkalmas tirozint tartalmazó ITAM molekulaszakaszok találhatók.
A B-sejt receptor (BCR) tehát molekulakomplex, amely jelfelismerõ és jeltovábbító alegységbõl épül fel (5. ábra). Az immunrendszer sejtjeinek membránhoz kötött receptorai között – amint erre késõbb fény derült – ez nem egyedülálló.
Az MHC-molekulák
A három antigénfelismerõ struktúra között az a jellegzetes és lényeges különbség, hogy az antigént különféle módon ismerik fel. Az elsõként ismertetett BCR (éppúgy, mint az ellenanyag-molekula) az antigént, ill. annak valamelyik epitópját közvetlenül képes felismerni. A T-limfociták antigénreceptora azonban erre nem alkalmas. Ilyenkor a szervezetbe jutó antigént az ún. antigén-prezentáló (bemutató) sejtek (APC) veszik fel, amelyeket azután emésztõ vakuolumaikban megemésztenek. A lebontás után az antigénbõl származó fehérjék a sejtben egy különleges peptidreceptorhoz, a fõhisztokompatibilitási génkomplex (MHC) termékéhez kötõdnek és azzal komplexet képezve jelennek meg az APC membránján. A T-sejt kizárólag az MHC-molekula kötõhelyében elhelyezkedõ, antigén-eredetû peptidet (MHC)-peptid komplexet) ismeri fel.
Az MHC-molekula szerkezetével, az antigén prezentáció folyamatával és biológiai jelentõségével a Természet Világa 1994. 2. számában Rajnavölgyi Éva: Többszintû felismerés nélkül nincs immunválasz címû írásában foglalkoztunk részletesen, ezért ezekre itt nem térünk ki. Röviden ismertetjük viszont az antigénfeldolgozás eredményeképpen képzõdõ peptideket megkötõ és azokat a T-sejt receptoroknak bemutató molekulákat, az MHC-génkomplex termékeit.
Az antigénbõl származó peptideket bemutató sejtfelszíni molekulák képzõdését az MHC-I és az MHC-II osztályba tartozó gének szabályozzák, termékeik az MHC-I-nek, ill. MHC-II-nek nevezett fehérjék. Az MHC-I glikoproteidek a szervezet szinte valamennyi magvas sejtjének membránján jelen vannak, míg az MHC-II membránfehérjék elsõsorban az immunrendszer sejtjein találhatók. Az MHC-molekulákra – éppúgy, mint a B- és T-sejtek antigénreceptoraira – rendkívül nagy szerkezeti sokféleség jellemzõ. Ez a sokféle antigén felismerését lehetõvé tevõ polimorfizmus azonban az MHC-molekulák, illetve a B- és T-sejt receptorok esetében másként alakul ki. Ez utóbbiak szerkezeti sokfélesége (amint erre a BCR esetében utalunk is) a sejt fejlõdése során lezajló génátrendezõdés és szomatikus mutációk eredménye. Az MHC-molekulák polimorfizmusa két más tényezõre vezethetõ vissza. Az egyik a poligenitás, ami azt jelenti, hogy az MHC-komplex I, ill. II régióiban több gén helyezkedik el; a másik pedig a genetikai polimorfizmus, ami annak tulajdonítható, hogy az MHC-I és -II osztályba tartozó géneknek számos különbözõ allélja van, amelyek az utódokban a mendeli öröklõdésmenetet követve, kodominánsan öröklõdnek.
Mindez azt is jelenti, hogy amíg a B- és T-sejtek antigénfelismerõ receptorkészlete az egyed élete folyamán a szomatikus génátrendezõdés miatt állandóan újratermelõdik, következésképpen változik, addig a szülõktõl örökölt és ugyancsak nagyfokú polimorfizmust mutató MHC mintázat az egyed sejtjeire jellemzõ és az egyed élete folyamán változatlan.
Jóllehet az MHC-I és -II membránfehérjék különbözõ alegységekbõl épülnek fel, térszerkezetileg sok tekintetben hasonlítanak egymáshoz, például abban, hogy mindkettõ felszínén az antigén feldolgozásának eredményeként peptidek megkötésére alkalmas “peptidkötõ hely” található (6. ábra).
6. ábra. Az MHC-I és MHC-II molekula peptidkötô
helyének szerkezete
(Az illusztrációk az Immunbiológia címû
könyv ábrái nyomán készültek)
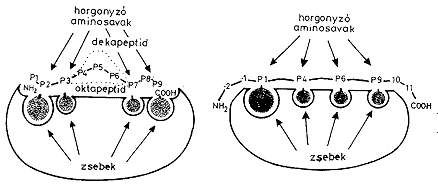
Az MCH-I és MHC-II molekulák felszínén tehát egyaránt kialakul az antigén feldolgozása során képzõdõ peptidek megkötésére alkalmas “zseb”, amely azonban nem monospecifikus. Ez azt jelenti, hogy egy adott allélt képviselõ MHC-molekula akár néhány száz eltérõ struktúrájú (de bizonyos szerkezeti jellegzetességet mutató) peptid megkötésére alkalmas. Ennek következtében az MHC-molekulák mint peptidreceptorok a szervezet saját fehérjéibõl származó peptideket éppúgy megkötik, mint a nem saját antigénekbõl származókat. Így a saját peptideket a T-sejteknek “bemutatva” biztosítják a saját fehérjék “eltûrését” (immuntolerancia); míg a nem saját struktúrákból származó peptidek T-effektor sejtek útján történõ felismerése a nem saját anyagok eltakarítását biztosítja a szervezetbõl.
Az MHC-I és MHC-II molekulák peptidkötõ helye szerkezeti hasonlóságuk ellenére funkcionálisan eltér egymástól. Az MHC-I molekulák kötõhelye “zárt”, azaz a kötõhely két végén olyan aminosavak találhatók, amelyek a peptidek N-, ill. C-terminális csoportjaival reagálnak. Emiatt a kötõhely csak meghatározott méretû peptideket fogad be. Az MHC-II molekulák peptidkötõ helyei “nyitottak”, azaz a peptiddel reagálni tudó aminosavak a kötõhely “fenekén”, annak hosszában helyezkednek el, így a peptid N-, ill. C-terminális végei mintegy kilógnak a zseb két szélén. Ennek következtében nagyobb és változó méretû peptidek kötõdhetnek az MHC-II molekulák peptidkötõ helyébe.
 |
| 7. ábra. A TC- és TH-sejtek MHC-függô antigénfelismerése közötti különbség |
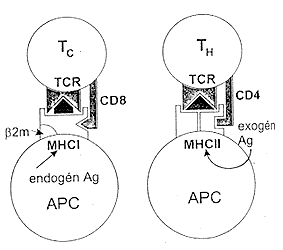
Természetesen felvetõdik a kérdés:
mi biztosítja a felismerés (következésképpen
az immunfolyamatok) fajlagosságát, ha az antigén prezentációjában
kulcsszerepet játszó MHC-molekulák nem tudják
a “saját” és “nem saját” eredetû peptideket
egymástól megkülönböztetni? Ez a fajlagosságot
biztosító differenciálás a
T-sejt receptorok funkciója: az antigénbõl származó
peptid az MHC-molekulák segítségével megjelenik
az APC felszínén és a T-sejt nagy specificitást
mutató antigénreceptora csak a szerkezeti sajátságainak
megfelelõ komplexet ismeri fel. Így a T-sejt antigénreceptora
az MHC-I molekulát és annak a “zárt” kötõhelyébõl
kitüremkedõ peptidszakaszát, ill. az MHC-II molekulát,
annak a “nyitott” kötõhelyében helyet foglaló
peptidet a kötõhelybõl kilógó részekkel
együtt “látja” (7. ábra).
(Zikernagel és Dougherty 1974-ben írták le az MHC-korlátozásnak nevezett törvényszerûséget, miszerint a T-sejtek az antigént csak az MHC-molekula társaságában képesek felismerni és ennek az immunológiai “alaptörvény”-nek a bizonyításáért 1996-ban Nobel-díjat kaptak. A témával folyóiratunk 1997. 1. száma foglalkozott: Falus András: “A kettõs felismerés. A T-limfociták jellegzetes látásmódja”.)
A T-sejt receptorkomplex
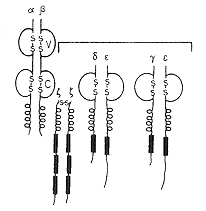
A T-sejt antigénfelismerõ receptora – hasonlóan a BCR-hez – a MIRR receptorcsaládba tartozik (8. ábra). Molekulakomplex, amelyben más-más alegység képviseli a felismerõ- és információt továbbító funkciót. A komplex egy kétláncos, négy doménbõl álló, nagy változatosságot mutató antigénfelismerõ egységbõl (a tulajdonképpeni TCR) és az ehhez kapcsolt CD3-komplexnek nevezett, nem variábilis fehérjébõl áll. A BCR-rel való hasonlóság abban is megnyilvánul, hogy mindkét receptorkomplex komponensei az Ig-szuperfamíliába tartoznak. Jellegzetes különbség viszont, hogy míg a BCR szolubilis formában (ellenanyag) is megjelenik, addig a TCR-nek csak membránhoz kötött változata ismert.
 |
| 8. ábra. A TRC-komplex szerkezete |
A TCR antigénfelismerõ egysége, amely egy meghatározott antigén eredetû peptid és MHC-molekula komplexével képes kölcsönhatásba kerülni, két polipeptidláncból épül fel. A receptorok túlnyomó többsége (ab-TCR) diszulfid híddal összekapcsolt a- és b-láncból álló heterodimer (kis hányaduk g- és d-nak nevezett láncokból áll, ezekre itt nem térünk ki), melyeknek a membrántól távolabb esõ, variábilis doménjei alkotják az antigénkötõ helyet. A TCR polimorfizmusa – hasonlóan a BCR-nél leírtakhoz – a T-sejt egyedfejlõdése során, szomatikus génátrendezõdés eredményeként alakul ki. A receptor transzmembrán szakaszának töltésviszonyai lehetõvé teszik a TCR-ral komplexet alkotó CD3-molekulákkal való kapcsolatot, citoplazmatikus szakasza (ugyancsak hasonlóan a BCR-hez) közvetlen jelátvitelre alkalmatlan. A jelátvitel a TCR esetében is a molekulához asszociált, jelátvitelre alkalmas szerkezetû molekulák funkciója, melyek sok hasonlóságot mutatnak a BCR-komplex Iga- és Igb-láncaival.
A CD3-komplex (8. ábra) három, nem kovalens módon kapcsolódó polipeptid láncból (g, d és e) és a hozzájuk kötõdõ dimer molekulákból épül fel (z,z-h). A g, d és e láncok egy N-terminális Ig-típusú extracelluláris domént, egy transzmembránt, ill. egy citoplazmatikus részt tartalmaznak. A transzmembrán szakaszok negatív töltése teszi lehetõvé az ab-heterodimérrel való funkcionális kölcsönhatás kialakulását. A citoplazmatikus részek meglehetõsen hosszúak, ITAM-motívumokat tartalmaznak, így jelátviteli folyamatokban való részvételre alkalmasak. Az egymáshoz hasonló struktúrájú z és h láncok a g, d és e láncoktól eltérõ szerkezetûek, de mindkettõben találhatók a jelátviteli mechanizmusok szempontjából fontos ITAM-motívumok. (Érdemes megjegyezni, hogy a z lánc más receptor-komplexben, így az FcgRIII-ban is megtalálható.)
| Természet Világa, | 129. évf. 11. sz. 1998. november, 482–486. o. https://www.kfki.hu/chemonet/TermVil/ https://www.ch.bme.hu/chemonet/TermVil/ |