A szúnyogok által terjesztett fertőző megbetegedések száma emelkedő tendenciát mutat a Földön, amit az elterjedési területük folyamatos bővülése is kísér. A trend hátterében a klímaváltozás hatását feltételezik a kutatók. Európában terjedőben lévő szúnyogok által terjesztett megbetegedés például a hazánkban is egyre nagyobb esetszámban előforduló Nyugat-nílusi láz. A klímaváltozáson kívül egyéb emberi hatások, így a földhasználat változása is hozzájárulhat a szúnyogok által terjesztett megbetegedések számának emelkedéséhez.
Az emberiség számára negatív trendek mellett az is aggodalomra ad okot, hogy jelenleg is komoly egészségügyi problémát jelentenek ezek a megbetegedések a trópusi és szubtrópusi területek lakossága számára. Csak maga a malária évente több, mint egymillió gyermek életét oltja ki, főként a Szaharától délre elterülő Afrikában. Magától értetődő, hogy számos kutatás igyekszik megérteni a szúnyogok által okozott veszélyek természetét, várható trendjeit és megtalálni a lehetséges védekezési módokat. A védekezés nem lehetséges a szúnyogok viselkedését, évszakos aktivitását, élőhelyválasztását meghatározó tényezők megértése nélkül. A kutatási célból vagy szúnyogkontrol miatt működtetett különféle csapdák hatékony működése is a szúnyogokra vonzást gyakorló fizikai és kémiai hatások ismeretét tételezi fel. Érdemes külön tárgyalni a szúnyoglárvák és az imágók viselkedési mintázatainak néhány aspektusát és az azokat befolyásoló faktorokat, hiszen eltérő, öröklött idegélettani mechanizmusok szükségesek a túléléshez a két, igen eltérő környezetben élő és anatómiájú fejlődési fázisban.
A ragadozókra adott viselkedési válaszok
Természetes kisvizekben pihenő Aedes és Culex szúnyoglárvákat megzavarva láthatjuk, hogy azok már a vízre vetülő árnyékunk hatására összerebbennek és a vízfelszín alá merülnek. Az Aedes aegypti esetében bizonyos nehézfémek (kadmium, króm és réz) gátolják a lárvák negatív fototaxisát. Ezzel szemben zavarásmentes esetben a Culex nemzetség fajainak, valamint az Aedes fuscus, Culiseta inornatus és Aedes curriei lárvái (és bábjai) pozitív fototaxist mutatnak, ami érthető, ha arra gondolunk, hogy normális esetben a víz/légkör határon tartózkodnak légzésük biztosítása végett. Sih (1986) igazolta, hogy a különböző szúnyog fajok lárváinak a potenciális ragadozókra adott válasza öröklött és fajspecifikus. Kisvizekben a hátonúszó poloskafajok (Notonecta nemzetség) számára a szúnyoglárvák könnyen elérhető táplálékot jelentenek. A szerző Notonecta undulata poloskát juttatott Culex pipiens és Aedes aegypti lárvatenyészetekbe (1. ábra).


1. ábra. Culex nemzetségbe tartozó szúnyoglárvák (felül) (Forrás: msmosquito.com)
és Notonecta undulata hátonúszó poloska (alul) (Forrás: britannica.com)
Ismert, hogy az előző faj lárvái gyakran fordulnak elő olyan természetes vizekben, ahol a hátonúszó poloska is előfordul. Ezzel ellentétben a második szúnyogfaj lárvái meleg, gyakran igen kis vizekben élnek, amit a hátonúszó poloskák nem kolonizálnak. Sih (1986) fő megfigyelései a következők voltak: 1) mindkét faj lárvái kevesebb mozgással és a tartózkodási helyük változtatásával válaszoltak a ragadozó jelenlétére, 2) a Culex-lárvák ritkábban estek áldozatul a ragadozóknak, 3) a Culex-lárvák az első pontban meghatározott elkerülési stratégiákat nagyobb mértékben mutatták, mint a másik faj lárvái, 4) a Culex-lárvák által adott magatartási válasz arányos volt a ragadozók számával, és 5) egyedül a Culex-lárvák mutattak predátorellenes viselkedést olyan vízmintákban, amelyekben előzőleg a poloskák fajtársaikat fogyasztották. A szerző azt a következtetést vonta le, hogy ameddig az Aedes-lárvák csak a ragadozó okozta direkt zavarásra adtak viselkedési választ (ez közös öröklött válaszreakció lehet több szúnyogfaj esetén), a Culex-lárvák az aktuális predációs nyomással kapcsolatos, annak mértékét jelző kémiai faktorok érzékelésére is képesek és arra adekvát elkerülési stratégiával válaszolnak, ami alacsonyabb halandóságukat magyarázza ilyen környezetben.
Fototaxis és táplálékkeresés
Tapasztalati tény, hogy más repülő rovarokhoz hasonlóan a szúnyogok imágóit kifejezetten vonzza a mesterséges világítótestek fehér fénye. A vonzás mértéke azonban fajonként változó. Wilton (1981) Culex pipiens pipiens és Culex pipiens quinquefasciatus nőstényeinek tesztpopulációit elengedve és fénycsapdával ismét foglyul ejtve azt találta, hogy az első alfajt vonzotta nagyobb mértékben a mesterséges fény, a másodikat kevésbé. A szerző a két alfaj hibridjeit is létrehozta, és azt tapasztalta, hogy ezek visszafogási aránya a fénycsapdával a két alfaj közötti értéket mutatott. Mindenesetre a fénycsapdák hatékonyságához kétség nem férhet (2. ábra).
2. ábra. Fénycsapda működés közben
A legtöbb szúnyogfaj nőstény egyede a melegvérű állatok és az ember vérét fogyasztja, de vannak kétéltűek vérét szívó fajok is (például Culex martinii). A gazdaállatok felkeresése és az eredményes vérszívás összetett alkalmazkodást és komplex anatómiai és viselkedési válaszokat követelt meg az evolúció során a szúnyogoktól, aminek vezérlésében alapvető szerepe van a szúnyogok szaglásának. A szúnyogok szagérzékelése nem csak táplálékkereső magatartásukat határozza meg alapvetően, de szexuális és szaporodóhely-találási képességüknek is fontos letéteményese (3. ábra).
:max_bytes(150000):strip_icc()/184940553-56a709945f9b58b7d0e631a9.jpg)
3. ábra. Emberi testfelszínen táplálkozó nőstény szúnyog (Forrás: thespruce.com)
A szén-dioxid-koncentráció érzékelése minden esetben központi helyet foglal el a gazdaszervezet lokalizációjában és gyakorlatilag minden szúnyogfaj számára fontos jelzés (tudniillik az állatok szén-dioxidot lélegeznek ki; a melegvérű állatok fokozott anyagcseréjük miatt többet, mint a hüllők vagy a kétéltűek). Emellett számos, a vért „szolgáltató” állatfajra jellemző szignált is érzékelnek a szúnyogok, ami vérforrástól függően eltérő és fajspecifikus lehet. Például, a különböző zsírsavak és a tejsav, amik az emberi izzadmányban különösen nagy mennyiségben fordulnak elő, érzékelhetők és vonzók számos, kifejezetten vagy alkalmilag embervért fogyasztó szúnyog számára. Az 1-oktén-3-ol (oktenol) és a széndioxid együttese a legfontosabb vonzerő az embervért kedvelő szúnyogok számára. Nemcsak az ember, de például a szarvasmarhák is kibocsátják az oktenolt (4. ábra).
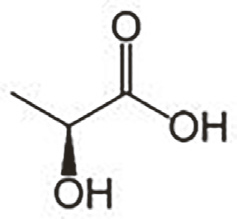

4. ábra. A tejsav (felül) és az 1-oktén-3-ol (R)-izomer
szerkezeti vázai (alul)
A maláriaszúnyogok közül az Anopheles gambiae – a Plasmodium falciparum, a malaria tropica okozójának legjelentősebb terjesztője – esetében kimutatták, hogy a csak a nőstény szúnyogokban termelődő AgOr1 proteinnek alapvető jelentősége van az emberi izzadságot érzékelni képes receptor kifejezésében. A receptorok fehérjéit kódoló génszakaszok jobb megismerésétől remélik a maláriaszúnyogok elleni védekezés egyik lehetséges, specifikus módjának kifejlesztését. Itt megjegyzendő, hogy egyes csapdafajták nem csak a fény, hanem a szén-dioxid vagy egyes vegyületek szúnyogokra kifejtett vonzó hatását is kihasználva működnek. Más anyagok, mint például az urzolsav-tartalmú csodatölcsér (Mirabilis jalapa), a rozmaringsavat nagy mennyiségben tartalmazó rozmaring (Rosmarinus officinalis) és az oleánsavat, urzolsavat és rozmaringsavat egyaránt tartalmazó tulsi (Ocimum tenuiflorum) kivonatai erős taszító hatást fejtenek ki a szúnyogokra.
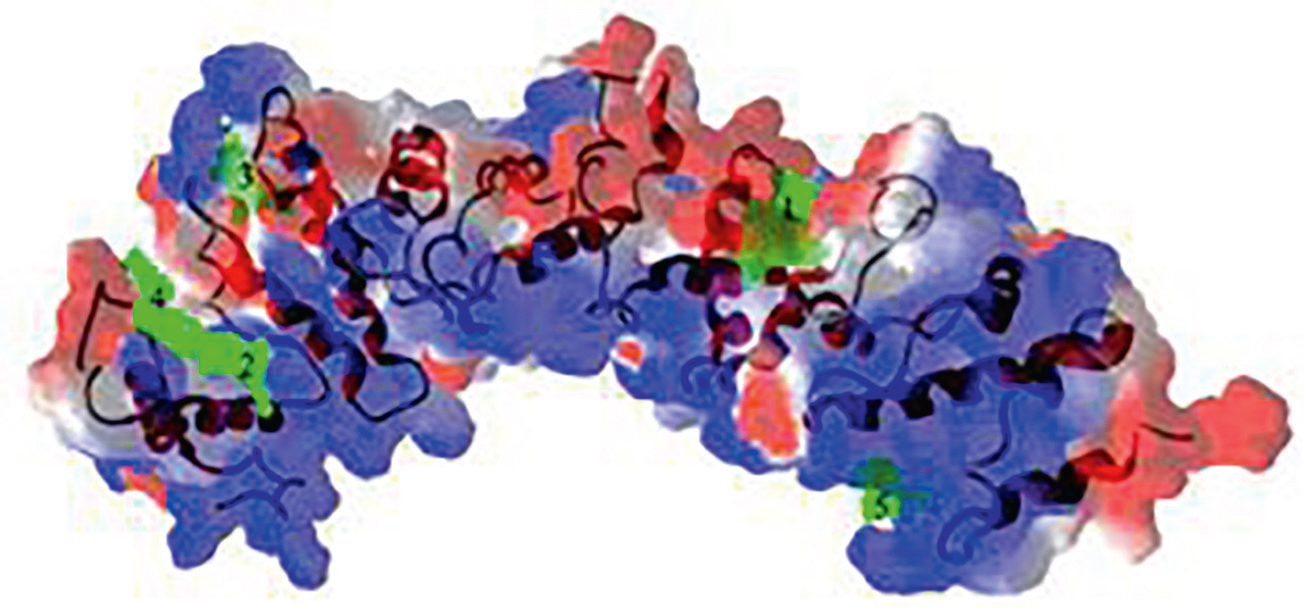
5. ábra. Az AgOR1-protein háromdimenziós képe
(Akash Deep Biswas értekezése nyomán, 2015)
Reproduktív viselkedés
A szaglásnak szerepe van a nőstények esetén a peterakás kiváltásában és helyének kiválasztásában is. A vízben oldott tejsavófehérje (lactalbumin), valamint bizonyos bakteriális termékek (például neomycin) attraktív hatással vannak a nőstény szúnyogokra. Ugyancsak bizonyítást nyert, hogy a 21 C-számú zsírsav-észtereknek erős hatása van a Aedes aegypti és Aedes albopictus szúnyogok peterakási aktivitására. Egyes hosszú láncú zsírsavészterek taszító (repellens), mások vonzó (attraktáns) hatással rendelkeznek. A jelenség hátterében az állhathat, hogy a lárvák fejlődéséhez szükséges környezetnek megfelelő mikrobiális közösséggel kell rendelkeznie. A fent említett vegyületek, mint indikátorok, jelentenek a víz állapotáról, jellegéről és az elérhető táplálékforrások jelenlétéről. Sumba és munkatársai (2007) természetes vízmintákat gyűjtöttek olyan helyekről, ahol szúnyoglárvák előfordultak, majd laboratóriumi körülmények között a vízminták egy részét módosították szűrt víz, autoklávozott (csírátlanított) talajminta vagy baktériumtenyészetek hozzáadásával. Azt találták, hogy a nőstény szúnyogok előszeretettel választották a természetes vízmintákat és kerülték a módosított és mesterséges vizeket.
A szagláson túl, hangérzékelés is fontos szerepet játszik a szúnyogok párkeresésében. Az emberi fülnek (nem véletlenül) oly idegesítő nőstény szúnyog zümmögés fontos párzásra késztető jel a hím szúnyogok számára. A hatás persze kölcsönös, mivel mindkét fél számára fontos szerepe van a „násztánc” során ezeknek a hangfrekvenciáknak. A szúnyogok „füle” a Johnston-féle szerv, ami akár 2000 Hz-es hangfrekvencia észlelésére is képes a felső rezgéstartományban. Az egymás jelenlétének felismerését elősegítő hangok kiadásának nagyon fontos szerepe van a szúnyogpopulációk fennmaradásában, mivel lényeges szempont, hogy egy adott életkorban termékenyüljenek meg az amúgy nem hosszú életű nőstény szúnyogok. A bábból kikerülő nőstények megtermékenyíthetősége erősen függ koruktól. Edman és munkatársai (1972) Aedes taeniorhynchus nőstény egyedeit vizsgálva például azt találták, hogy a legtöbb nőstény egyed 30-40 órával a bábból való kikelése után termékenyült meg alig valamivel napnyugta után. Más szerzők kimutatták, hogy bár a nőstény és hím Aedes aegypti hangkibocsátási alapfrekvenciája 400 és 600 Hz, a párzási frekvencia eléri a közösen hallatott 1200 Hz értéket. Hasonló ivari eltérés ismert a hím és nőstény szúnyogok által kibocsátott hangfrekvenciák tekintetében az Aedes aegypiti esetében is. Érdekes, hogy bár az azonos területen előforduló Anopheles funestus és Anopheles gambiae szúnyogok nőstényei által kibocsátott hangfrekvencia nagyon hasonló (516-497 Hz), ez nem zavarja meg a szúnyogok nászát.
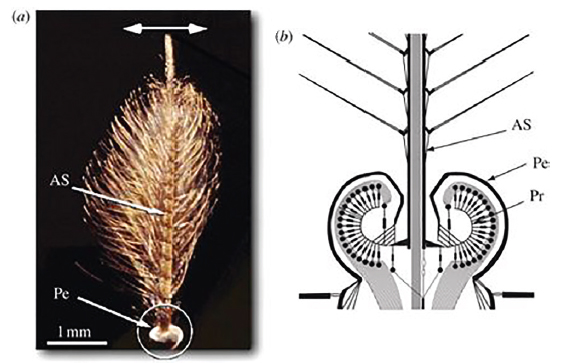
6. ábra. Egy hím szúnyog csápja és a Johnston-féle szerv a csáp alapi részén (AS: csápostor, Pe: kocsány, Pr: szkolopidiumok)
(Forrás: royalsocietypublishing.org)
Rajzás, tájékozódás
A rajzás a szaporodási viselkedéshez szorosan kapcsolódik. Charlwood és Jones (1980) az Anopheles gambiae fajjal kapcsolatban megfigyelték, hogy a faj hímjei a mangrove mocsarak szélén előforduló tisztásokon rajzanak előszeretettel, mely viselkedés mesterséges tisztások létesítésével is kiváltható. A nőstények a hímek alkotta rajba repülnek be párosodás céljából. A szerzők kísérletet is végeztek a faj hímjeivel és nőstényeivel. Elsötétített laboratóriumukban egy fekete területet létesítettek a padlón. Azt tapasztalták, hogy miközben a hímek szabályosan rajzottak a sötét rész felett, addig a nőstények csak rövid időre kerültek a terület fölé, majd onnan gyorsan visszafordultak. Azt a következtetést vonták le megfigyeléseikből, hogy amíg a hímek vizuálisan keresnek „arénákat” (kis tisztásokat) a párzáshoz és annak megtörténtéig ott is maradnak, addig a nőstények keresik ezeket a helyeket addig, ameddig meg nem találják a hímek által alkotott rajokat (7. ábra).
7. ábra. Nappali szúnyograjzás Alaszkában 2013 júliusában
A rajzást gyakran megzavarhatják a ragadozók, ugyanakkor a párzás sikeressége populációs szempontból – más rajzó fajokhoz hasonlóan – lényegesen jobb, mintha a faj nem rajban párzana. Yuval és Bouskila (1993) megfigyelték, hogy az Anopheles freeborni szaporodási sikere a párzások száma/a szitakötők okozta zsákmányolások aránya alapján jobb hatásfokú erdőfoltokban, mint a nagy, nyílt területeken, tisztásokon. Charlwood és munkatársai (2003) az Anopheles funestus napnyugta után közvetlenül kezdődő rajzását vizsgálva azt találták, hogy a rajzás kezdete után az első párok 11 perccel voltak megfigyelhetők, valamint számuk fordítottan arányos volt a rajzás megkezdése óta eltelt idővel. Azt is észrevették, hogy a nőstények túlnyomó többsége csak a párosodást követően táplálkozott.
A felületek polarizációs tulajdonságai fontos szerepet játszanak a megfelelő petézőhelyek kiválasztásában számos vizekben fejlődő rovar esetében. Bernáth és munkatársainak (2008) vizsgálata szerint azonban az Aedes aegypti nőstényei nem rendelkeznek polarotaxissal, ami azzal magyarázható, hogy az általuk keresett kisvizek nem azonosíthatók ilyen módon. Ezzel szemben, a nagyobb vizekben (tavak, folyók) szaporodó árvaszúnyog-fajok (család: Chironomidae) nőstényei érzékelik a vizek polarizációs jellegzetességeit, ahogy a kérészek (család: Ephemeridae) nőstényei is. A különbség hátterében az áll, hogy ameddig az árvaszúnyog- és kérészlárvák az iszapban fejlődve általában elkerülik a ragadozókat, így kolonizálhatják a nagyobb vizeket is, a szúnyogok lárvái a vízfelszínhez vagy az alámerült vegetációhoz kötődnek, így igen könnyű célpontjai a vízben szabadon mozgó, vizuális predátoroknak.
Napszakos aktivitás
Emberi környezetben nagy jelentősége lehet a szúnyogok által terjesztett fertőzések átvitelének esélye szempontjából, hogy egy adott szúnyogfaj a nappali vagy az éjszakai órákat részesíti-e előnyben táplálkozás szempontjából, illetve, hogy a házon/lakáson kívül vagy a belterekben is mutat táplálékkereső aktivitást. Ismert például, hogy egyes maláriaszúnyogok – mint amilyen például a dél-amerikai Anopheles darlingi vagy az Anopheles deaneorum sokkal gyakrabban csíp az épületekben, mint más maláriaszúnyog fajok nőstényei. Ha lakott területen együtt fordulnak elő, akkor az Aedes aegypti a beltérben, az Aedes albopictus pedig a házon kívül szerzett szúnyogcsípések túlnyomó részéért lehet felelős. A két Aedes-faj beltéri csípési aktivitása is szabályos napi ritmust mutat, mivel jóval nagyobb számban szenvednek el az emberek csípést a délelőtti órákban, mint kora délután. Ezzel szemben a legtöbb hazai szúnyogfaj a naplemente utáni félhomályban a legaktívabb. A Culex tarsalis esetében például azt találták, hogy naplemente után 2 órával éri el aktivitásának csúcsát. Tapasztalat szerint napnyugta után, ahogy hűl a levegő hőmérséklete, a fénycsapdával gyűjthető szúnyogok száma egyre csökken éjfél felé közeledve (8. ábra).
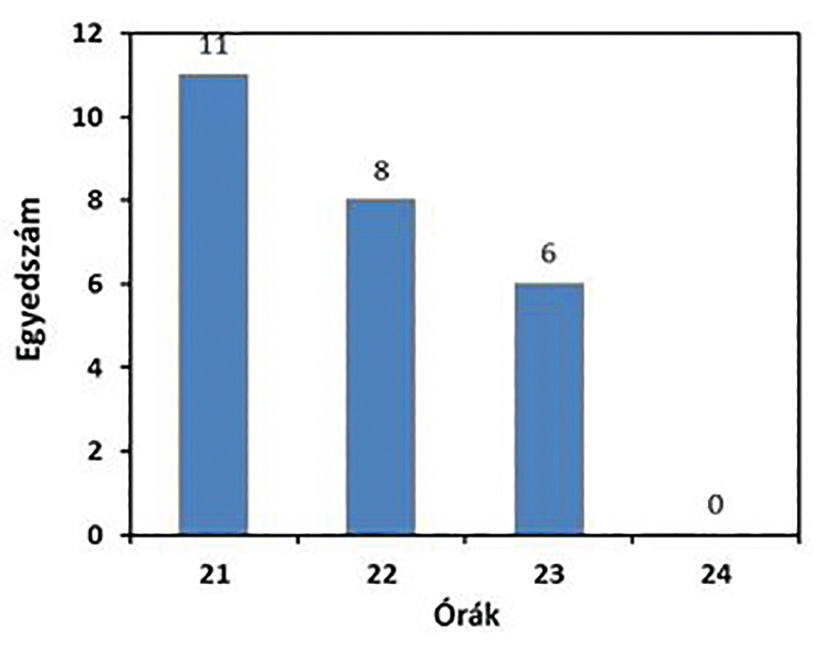
8. ábra. Egy Nagyharsányban végzett fénycsapdás gyűjtés egy estéjére vonatkozó, óránként gyűjtött szúnyogok egyedszáma
(Trájer Attila János és Tánczos Balázs gyűjtése alapján, 2014)
Repülés, terjedés
A szúnyogok repülési képessége nem a legjobb a rovarok világában, de az is megjegyzendő, hogy a szúnyogok terjedésének lehetőségét erősen behatárolja alacsony túlélési idejük (a mindennapi túlélési esély az Aedes notoscriptus esetében pl. 77-79%. Jelölési–elengedési–befogási (mark–release–recapture) tanulmányok alapján ismert, hogy a Culex tarsalis egy nap alatt mindössze nagyjából 500 méter megtételére képes saját repülési képességére támaszkodva. A Culex quinquefasciatus nőstények esetében hasonló vizsgálatokkal 0,6-1 km/nap terjedési értékeket kaptak. Egy, az Aedes aegypti nőstényeivel végzett kísérlet 15 nappal az elengedést követően a fluoreszceinnel jelölt szúnyogok mindegyikét 200 m sugáron belül találta és az átlagos eltávolodás a forrástól mindössze 78 méter volt. Egy másik tanulmány 160 méter maximális eltávolodást és 56, valamint 35 méter átlagos szóródási távolságot mért az Aedes aegypti nőstényei és hímjei esetében a kibocsátási helytől. Ez a feltűnő, nemek közötti különbség talán arra utalhat, hogy a faj terjedése szempontjából a nőstények repülési távolságának nagyobb jelentősége van, mint a hímek szóródási képességének, hiszen, mint már említettük, a nőstény szúnyogok már a kikelésüktől számított néhány napon belül általában megtermékenyülnek, meghatározva fajuk természetes terjedési sebességét (például az Anopheles gambiae esetében a második napon volt a legnagyobb a nőstény szúnyog megtermékenyülésének esélye (9. ábra).

9. ábra. Az Anopheles gambiae maláriaszúnyog-faj frissen vért szívó nősténye
(Forrás: igtrcn.org)
A szúnyogok terjedését azonban más, fizikai, biológiai és antropogén faktorok is erőteljesen befolyásolják. Felmerül az a lehetőség is, hogy a délről érkező meleg légtömegeknek szerepe lehet a szúnyogok szaltatórikus, nagy távolságokon átívelő, dél-észak irányú terjedésében. Sellers (1989) megállapította, hogy az 1972-ben Québec és Connecticut államokban észlelt keleti ló encephalitis (agyvelőgyulladás) járvány kialakulásában szerepe volt a déli szeleknek. A trópusi ciklonok kiváltképp alkalmasak arra, hogy nagy magasságba emeljék és nedves légtömegükben akár több száz kilométeres távolságokon át épségben szállítsák az elragadott rovarokat. Arra is volt példa, hogy fordított irányban, északról délre szállított a szél szúnyogokat. Ez történt például 1995-ben és 1998-ban, amikor Új-Guineából egyenesen Ausztráliába transzportálták az erős szelek az élő és a Japán-B-encephalitis vírussal fertőzött szúnyog egyedeket. A szelek transzport szerepe azért rendkívül fontos, mert a gyenge repülő képességű, a sós vízben lárvaként terjedni nem képes és a direkt napsugárzással szemben védtelen rovarok olyan távolságokat képesen passzív módon megtenni, amire máskülönben soha nem lennének képesen természetes úton. Akad még egy, bár egyelőre kevéssé dokumentált lehetőség, miszerint lárvák és peték esetleg a vízimadarak tollazatához, illetve a madarak lábához tapadva is képesek lehetnek nagy utak megtételére. Egyelőre az látszik biztosnak, hogy gombák, magvak és egyes vízi gerinctelenek a madarak testfelületére tapadva is nagy távolságok megtételére képesek. Ma már egészen nyilvánvaló, de nehezen modellezhető az emberi hatások szerepe is a szúnyog fajok terjedésében. Egy példát említve, a fontos betegségterjesztő ázsiai tigrisszúnyog a szerencsebambuszként eladott, vízkultúrákban tartott és szállított Dracaena marginata-val került Floridába a távol-keleti eredeti előfordulási területéről. Az ázsiai tigrisszúnyog terjedéséről és ökológiai igényeiről bővebben olvashatnak a Természet Világa 2017 áprilisában megjelent, 148/4. számában.
Nyugalmi állapot, telelés
Mérsékelt övi körülmények között a téli fagy miatt egyes szúnyog fajok kizárólag pete vagy lárva, mások részben vagy teljesen imágó formában telelnek át. Az Egyesült Királyságban végzett vizsgálataik során Sulaiman and Service (2007) azt tapasztalták, hogy a Culex pipiens nőstény egyedei már júliusban vagy augusztusban megkezdik elvonulásukat téli szálláshelyeikre, maximális számukat pedig októberben vagy novemberben érik el ezeken a helyeken, amik lehetnek természetes vagy mesterséges üregek, fűtetlen, de hőszennyezett épületrészek is. Az első fagyok beálltával a szúnyogok távolabb húzódnak a bejárattól. A szúnyogok magas téli halandósága mögött a pókok okozta predáció, kórokozó gombák okozta elhullás és feltehetően a glikogén tartalékok idő előtti felélése is szerepet játszhat. Megfigyelték, hogy a telelő Culex pipiens szúnyogok gyakran váltogatják pozíciójukat a telelőhelyen belül és a kora hibernáció idején (augusztustól októberig) legalább 12 napig tartó folyamatos megvilágítással lehetett a nőstények táplálékkereső magatartását kiváltani, ami a hónapok előrehaladtával egyre csökkent. Ez rávilágít arra, milyen fontos moduláló szerepet tölt a téli hibernáció kiváltásában a nappalok hosszának éves ritmusa, valamint az átlaghőmérséklet. A svédországi, áttelelő Culex pipiens és Culex torrentium egyedek testében talált fruktóz arra utalhat, hogy a nőstény szúnyogok áttelelésének kulcsa a hibernáció előtti nektárfogyasztásuk, ami extra tápanyagforrást jelent számukra az ínséges téli hónapok során (10. ábra).

10. ábra. Barlang falán pihenő szúnyog
(Forrás: cambriancavingcouncil.org.uk)
Természetesen, a jelen írás terjedelemi korlátjai miatt sem vállalkozhatott arra, hogy a szúnyogok viselkedésének repertoárját részletesen, értekezés-szerűen ismertesse és az ezekkel kapcsolatos hipotéziseket bemutassa. Mindössze a főbb szempontok, a jelentősebb faktorok és viselkedési jellegzetességek ábrázolására volt lehetőség a teljesség igénye nélkül. Talán ennyiből is érzékelhető, hogy a mégoly kis idegrendszerrel bíró lények, mint a szúnyogok esetében is komplex öröklött viselkedési válaszok segítik a szúnyog populációk hosszú távú fennmaradását. A szúnyogok gyűjtése, elrettentése, illetve általános kontrolja is sok elemében megkívánja a szúnyogok viselkedésének ismeretét és jelenleg is aktív kutatások tárgyát képezi.
IRODALOM
[1] Bernáth, B., Horváth, G., Gál, J., Fekete, G., & Meyer-Rochow, V. B. (2008). Polarized light and oviposition site selection in the yellow fever mosquito: no evidence for positive polarotaxis in Aedes aegypti. Vision Research, 48(13), 1449-1455.
[2] Charlwood, J. D., Thompson, R., & Madsen, H. (2003). Observations on the swarming and mating behaviour of Anopheles funestus from southern Mozambique. Malaria Journal, 2(1), 2.
[3] Charlwood, J. D., & Jones, M. D. R. (1980). Mating in the mosquito, Anopheles gambiae sl. Physiological Entomology, 5(4), 315-320.
[4] Edman, J. D., Haeger, J. S., Bidlingmayer, W. L., Dow, R. P., Nayar, J. K., & Provost, M. W. (1972). Sexual behavior of mosquitoes. 4. Field observations on mating and insemination of marked broods of Aedes taeniorhynchus. Annals of the Entomological Society of America, 65(4), 848-852.
[5] Sellers, R. F. (1989). Eastern equine encephalitis in Quebec and Connecticut, 1972: introduction by infected mosquitoes on the wind? Canadian Journal of Veterinary Research, 53(1), 76.
[6] Sih, A. (1986). Antipredator responses and the perception of danger by mosquito larvae. Ecology, 67(2), 434-441.
[7] Sulaiman, S., & Service, M. W. (1983). Studies on hibernating populations of the mosquito Culex pipiens L. in southern and northern England. Journal of Natural History, 17(6), 849-857.
[8] Sumba, L. A., Guda, T. O., Deng, A. L., Hassanali, A., Beier, J. C., & Knols, B. G. (2004). Mediation of oviposition site selection in the African malaria mosquito Anopheles gambiae (Diptera: Culicidae) by semiochemicals of microbial origin. International Journal of Tropical Insect Science, 24(03), 260-265.
[9] Yuval, B., & Bouskila, A. (1993). Temporal dynamics of mating and predation in mosquito swarms. Oecologia, 95(1), 65-69.
[10] Wilton, D. P. (1981). Light-trap response and the DV/D ratio in the Culex pipiens complex (Diptera: Culicidae). Journal of Medical Entomology, 18(4), 284-288.
A cikk a Természet Világa 2019. júniusi számában (150. évf. 6. sz.) jelent meg.

